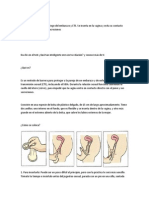
También podría gustarte
- Detección y diagnóstico de la peste porcina africana: Manual para veterinariosDe EverandDetección y diagnóstico de la peste porcina africana: Manual para veterinariosAún no hay calificaciones
- Sincronización de CeloDocumento26 páginasSincronización de Celopepe100% (1)
- Comportamiento productivo y reproductivo en caprinos en trópico secoDe EverandComportamiento productivo y reproductivo en caprinos en trópico secoCalificación: 1 de 5 estrellas1/5 (1)
- Castidad en Los NoviosDocumento8 páginasCastidad en Los NoviosParroquiaItatiVillaChicaAún no hay calificaciones
- Operaciones auxiliares en reproducción ganadera. AGAX0108De EverandOperaciones auxiliares en reproducción ganadera. AGAX0108Aún no hay calificaciones
- UF2165 - Manejo de animales reproductoresDe EverandUF2165 - Manejo de animales reproductoresAún no hay calificaciones
- Sincronización de Estro y Ovulación Diana AvilesDocumento19 páginasSincronización de Estro y Ovulación Diana AvilesCarlos Xavier RodriguezAún no hay calificaciones
- Marcadores Genéticos para El Mejoramiento de La Terneza de La Carne GENETICADocumento11 páginasMarcadores Genéticos para El Mejoramiento de La Terneza de La Carne GENETICAVenezuelaGanadera.comAún no hay calificaciones
- Informe ReproducciónDocumento14 páginasInforme ReproducciónVANESSA ALEXANDRA MURILLO LESMES100% (1)
- Calidad Seminal de TorosDocumento41 páginasCalidad Seminal de TorosErnestoRiascos100% (1)
- Ciclo EstralDocumento12 páginasCiclo EstralZabdiel Luna SoriaAún no hay calificaciones
- Constantes Fisiológicas de Los Animales DomésticosDocumento2 páginasConstantes Fisiológicas de Los Animales Domésticositzel Guadalupe PereydaAún no hay calificaciones
- Nutrición de Búfalos en Regiones TropicalesDocumento43 páginasNutrición de Búfalos en Regiones TropicalesAlejandro Londoño100% (17)
- Abdomen Agudo GinecológicoDocumento77 páginasAbdomen Agudo Ginecológicocarlospalominocruzad100% (1)
- Reproduccion Animal 3 - Ruben MoralesDocumento19 páginasReproduccion Animal 3 - Ruben MoralesRUBEN MORALES BETANCURAún no hay calificaciones
- INDICADORES Bienestar Vacas LecherasDocumento7 páginasINDICADORES Bienestar Vacas LecherasAlonso Antonio ArroyoAún no hay calificaciones
- Guía completa sobre métodos anticonceptivosDocumento3 páginasGuía completa sobre métodos anticonceptivosNhms CndrAún no hay calificaciones
- EquinosDocumento14 páginasEquinosManola Carrera CarbonelAún no hay calificaciones
- La Sexualidad en La Época ModernaDocumento7 páginasLa Sexualidad en La Época ModernaAle GuerraAún no hay calificaciones
- Urolitiasis en OvinosDocumento14 páginasUrolitiasis en Ovinosomarzito_mexflow5041100% (2)
- Parámetros Productivos y Reproductivos de Importancia Económica en Ganadería Bovina TropicalDocumento13 páginasParámetros Productivos y Reproductivos de Importancia Económica en Ganadería Bovina TropicalJose LuisAún no hay calificaciones
- Evaluación y Procesamiento de SemenDocumento6 páginasEvaluación y Procesamiento de SemenJigoku NekoAún no hay calificaciones
- Introducción a la producción ovina en México: situación actual, inventarios, razas y sistemas de producciónDocumento16 páginasIntroducción a la producción ovina en México: situación actual, inventarios, razas y sistemas de producciónMS-AAAún no hay calificaciones
- Parámetros Productivos y Reproductivos de Importancia Económica en Ganadería Bovina TropicalDocumento14 páginasParámetros Productivos y Reproductivos de Importancia Económica en Ganadería Bovina TropicalMaría Julia González QuesadaAún no hay calificaciones
- Sincronización de Estro BovinoDocumento8 páginasSincronización de Estro BovinoCarlos MartínezAún no hay calificaciones
- Hipertensión Pulmonar Arterial: (Ascitis) En Pollos De EngordaDe EverandHipertensión Pulmonar Arterial: (Ascitis) En Pollos De EngordaAún no hay calificaciones
- Enfermedades Mas Comunes en CerdosDocumento4 páginasEnfermedades Mas Comunes en CerdosAngel Alfonso Huerta Silva100% (2)
- Anestro en BovinosDocumento15 páginasAnestro en BovinosAnita Elizabeth Layza Mendiola50% (2)
- Ovinos y CaprinosDocumento23 páginasOvinos y CaprinosZeltha Marelens OhuraAún no hay calificaciones
- Condición Corporal en Bovinos Productores de CarneDocumento9 páginasCondición Corporal en Bovinos Productores de CarneJUAN MANUEL CABRALESAún no hay calificaciones
- Internado BovinoDocumento2 páginasInternado BovinoKary Isabel Fuenzalida MaripanguiAún no hay calificaciones
- Gallinas en Pastoreo para Producción de Huevo y La Engorda de Pollos y Guajolotes Es Una Alternativa ViableDocumento38 páginasGallinas en Pastoreo para Producción de Huevo y La Engorda de Pollos y Guajolotes Es Una Alternativa Viablenegro_pinolero9621Aún no hay calificaciones
- 1 HeredabilidadDocumento10 páginas1 Heredabilidadernesto luis crespin padillaAún no hay calificaciones
- Fisiología de La Reproducción de Los Animales DomésticosDocumento8 páginasFisiología de La Reproducción de Los Animales DomésticosElvis DugarteAún no hay calificaciones
- OvinosDocumento37 páginasOvinosNahuel Adrian VelezAún no hay calificaciones
- Anatomía y Fisiología Normal de La Reproducción en Felinos Hembra y MachoDocumento7 páginasAnatomía y Fisiología Normal de La Reproducción en Felinos Hembra y MachoCamilla MartinsAún no hay calificaciones
- Anatomia Glandula Mamaria Bovina Su Relacion Con La Produccion de Leche y La MastitisDocumento13 páginasAnatomia Glandula Mamaria Bovina Su Relacion Con La Produccion de Leche y La MastitisVenezuelaGanadera.comAún no hay calificaciones
- Eval - Poten - Repro - Macho Bovino PDFDocumento63 páginasEval - Poten - Repro - Macho Bovino PDFNICOLE BLANCOAún no hay calificaciones
- Coccidiosis en Bovinos.Documento5 páginasCoccidiosis en Bovinos.Sebastian ToroAún no hay calificaciones
- ANESTRODocumento5 páginasANESTRODiulan MéndezAún no hay calificaciones
- Diarrea NeonatalDocumento13 páginasDiarrea NeonatalCAPRINOS BAJA CALIFORNIA SUR, MEXICOAún no hay calificaciones
- Inseminación Artificial en Cerdos Y BovinosDocumento26 páginasInseminación Artificial en Cerdos Y Bovinosmoisito03100% (1)
- Manual Aves DiprodalDocumento22 páginasManual Aves DiprodalFamilia Inostroza InfanteAún no hay calificaciones
- Pubertad en Hembras BovinasDocumento11 páginasPubertad en Hembras BovinasFabian MartinezAún no hay calificaciones
- Neumonía Enzootica Porcina (A+)Documento16 páginasNeumonía Enzootica Porcina (A+)Pablo ValienteAún no hay calificaciones
- Alteraciones Aparato Locomotor BovinoDocumento12 páginasAlteraciones Aparato Locomotor BovinoFrancisco Córdoba Duque0% (1)
- Razas LecherasDocumento71 páginasRazas LecherasDamian Jimenez Mendez0% (3)
- Manejo del hato reproductor porcinoDocumento19 páginasManejo del hato reproductor porcinoMalu Vazquez CruzAún no hay calificaciones
- Dinámica folicular bovina: reclutamiento, selección y dominanciaDocumento28 páginasDinámica folicular bovina: reclutamiento, selección y dominanciaMvz Gabriel Gutierrez RodriguezAún no hay calificaciones
- Inseminacion Intrauterina ProfundaDocumento19 páginasInseminacion Intrauterina ProfundaYoannis Milovna Butron SolisAún no hay calificaciones
- 2020 Parametros Reproductivos EficienciaDocumento21 páginas2020 Parametros Reproductivos Eficiencialuis pablo SistiAún no hay calificaciones
- Ciclo EstralDocumento3 páginasCiclo EstralmassieuAún no hay calificaciones
- Sincronizacion de EstrosDocumento28 páginasSincronizacion de EstrosFernando Antonio Rojas ArteagaAún no hay calificaciones
- Condicion Corporal en BovinosDocumento4 páginasCondicion Corporal en BovinosDarwin NavarroAún no hay calificaciones
- Principales Enfermedades Que Afectan Al Ganado Productor de Leche Raza HolsteinDocumento17 páginasPrincipales Enfermedades Que Afectan Al Ganado Productor de Leche Raza HolsteinIrvin Tintaya MurilloAún no hay calificaciones
- Enfermedades VíricasDocumento20 páginasEnfermedades VíricasJohanSeSamaniegoAún no hay calificaciones
- Citologia Vaginal ExfoliativaDocumento19 páginasCitologia Vaginal ExfoliativaJuan-quii Ormaza ChicaAún no hay calificaciones
- Suplementacion Estrategica de Vacas Doble Proposito PartoDocumento18 páginasSuplementacion Estrategica de Vacas Doble Proposito PartoVenezuelaGanadera.comAún no hay calificaciones
- Ciclos reproductivos femeninosDocumento4 páginasCiclos reproductivos femeninosKimberly Capdevila100% (1)
- Teis de Brucelosis en Robore 2006Documento71 páginasTeis de Brucelosis en Robore 2006Eunice RojasAún no hay calificaciones
- Vitaminas en La Producción AnimalDocumento3 páginasVitaminas en La Producción AnimalRepositorio de la Biblioteca de la DEA-MAG-PYAún no hay calificaciones
- Parámetros Reproductivos en Bovinos Ciclo 2018-IDocumento5 páginasParámetros Reproductivos en Bovinos Ciclo 2018-IRuthy Sugey García SánchezAún no hay calificaciones
- Retencion Placentaria. ReproduccionDocumento5 páginasRetencion Placentaria. ReproduccionVenezuelaGanadera.comAún no hay calificaciones
- Zootecnia de OvinosDocumento46 páginasZootecnia de OvinosFrancisco44 likeAún no hay calificaciones
- Genética General y Aplicada. Unidad II.Documento22 páginasGenética General y Aplicada. Unidad II.Andres OrtegaAún no hay calificaciones
- Leguminosas espontaneas de posible valor forrajero en ColombiaDe EverandLeguminosas espontaneas de posible valor forrajero en ColombiaAún no hay calificaciones
- ¿Es La Neosporosis Un Problema A Considerar en El Búfalo de Agua (Bubalus Bubalis) ?Documento7 páginas¿Es La Neosporosis Un Problema A Considerar en El Búfalo de Agua (Bubalus Bubalis) ?VenezuelaGanadera.comAún no hay calificaciones
- Modelos Estadísticos de Evaluación Genética GENETICADocumento13 páginasModelos Estadísticos de Evaluación Genética GENETICAVenezuelaGanadera.comAún no hay calificaciones
- Genética Molecular para El Mejoramiento de Los Rasgos Reproductivos en Bovinos GENETICADocumento9 páginasGenética Molecular para El Mejoramiento de Los Rasgos Reproductivos en Bovinos GENETICAVenezuelaGanadera.comAún no hay calificaciones
- Innovadora Técnica de Lactoinducción para Mejorar La Productividad en La Ganadería Doble Propósito MEDICINADocumento9 páginasInnovadora Técnica de Lactoinducción para Mejorar La Productividad en La Ganadería Doble Propósito MEDICINAVenezuelaGanadera.comAún no hay calificaciones
- Estrategias Esenciales de Un Programa de Mejoramiento Genético Bovino (PMG) GENETICADocumento7 páginasEstrategias Esenciales de Un Programa de Mejoramiento Genético Bovino (PMG) GENETICAVenezuelaGanadera.com100% (1)
- Consideraciones para El Ajuste Del Modelo de Producción en El Sistema de Ganadería de Doble Propósito MEDICINADocumento7 páginasConsideraciones para El Ajuste Del Modelo de Producción en El Sistema de Ganadería de Doble Propósito MEDICINAVenezuelaGanadera.comAún no hay calificaciones
- Síndrome Abortivo y de Muerte PerinatalDocumento11 páginasSíndrome Abortivo y de Muerte PerinatalVenezuelaGanadera.comAún no hay calificaciones
- Logros y Desafíos en La Cría de Becerros Desde El Nacimiento Hasta El Destete en Ganaderías Doble PropósitoDocumento10 páginasLogros y Desafíos en La Cría de Becerros Desde El Nacimiento Hasta El Destete en Ganaderías Doble PropósitoVenezuelaGanadera.comAún no hay calificaciones
- Resistencia A Enfermedades en La Ganadería Doble Propósito, Un Enfoque Genético GENETICADocumento10 páginasResistencia A Enfermedades en La Ganadería Doble Propósito, Un Enfoque Genético GENETICAVenezuelaGanadera.comAún no hay calificaciones
- Los Beneficios Del Diagnóstico Molecular en La Ganadería Doble Propósito GENETICADocumento8 páginasLos Beneficios Del Diagnóstico Molecular en La Ganadería Doble Propósito GENETICAVenezuelaGanadera.comAún no hay calificaciones
- Enfermedades Heredables de Las Razas para Cruzamientos GENETICADocumento11 páginasEnfermedades Heredables de Las Razas para Cruzamientos GENETICAVenezuelaGanadera.com100% (1)
- Evaluación Genética de Reproductores - Logros y Desafíos GENETICADocumento20 páginasEvaluación Genética de Reproductores - Logros y Desafíos GENETICAVenezuelaGanadera.comAún no hay calificaciones
- Descarte, Vida Productiva y Selección en Rebaños Doble Propósito GENETICADocumento8 páginasDescarte, Vida Productiva y Selección en Rebaños Doble Propósito GENETICAVenezuelaGanadera.comAún no hay calificaciones
- El Germoplasma Criollo Imprescindible en La Ganadería Doble Propósito GENETICADocumento10 páginasEl Germoplasma Criollo Imprescindible en La Ganadería Doble Propósito GENETICAVenezuelaGanadera.comAún no hay calificaciones
- Desafíos para El Mejoramiento Genético de La Ganadería Doble Propósito GENETICADocumento11 páginasDesafíos para El Mejoramiento Genético de La Ganadería Doble Propósito GENETICAVenezuelaGanadera.comAún no hay calificaciones
- El Modelo Animal Simple - Una Metodología para Los Genetistas GENETICADocumento17 páginasEl Modelo Animal Simple - Una Metodología para Los Genetistas GENETICAVenezuelaGanadera.comAún no hay calificaciones
- Cruzamientos en La Produccion de Leche en El Tropico GENETICADocumento9 páginasCruzamientos en La Produccion de Leche en El Tropico GENETICAVenezuelaGanadera.comAún no hay calificaciones
- ¿Es La Neosporosis Un Problema A Considerar en El Búfalo de Agua (Bubalus Bubalis) ?Documento7 páginas¿Es La Neosporosis Un Problema A Considerar en El Búfalo de Agua (Bubalus Bubalis) ?VenezuelaGanadera.comAún no hay calificaciones
- Consideraciones para El Ajuste Del Modelo de Producción en El Sistema de Ganadería de Doble Propósito MEDICINADocumento7 páginasConsideraciones para El Ajuste Del Modelo de Producción en El Sistema de Ganadería de Doble Propósito MEDICINAVenezuelaGanadera.comAún no hay calificaciones
- Síndrome Abortivo y de Muerte PerinatalDocumento11 páginasSíndrome Abortivo y de Muerte PerinatalVenezuelaGanadera.comAún no hay calificaciones
- Modelos Estadísticos de Evaluación GenéticaDocumento13 páginasModelos Estadísticos de Evaluación GenéticaVenezuelaGanadera.comAún no hay calificaciones
- Logros y Desafíos en La Cría de Becerros Desde El Nacimiento Hasta El Destete en Ganaderías Doble PropósitoDocumento10 páginasLogros y Desafíos en La Cría de Becerros Desde El Nacimiento Hasta El Destete en Ganaderías Doble PropósitoVenezuelaGanadera.comAún no hay calificaciones
- ¿Es La Neosporosis Un Problema A Considerar en El Búfalo de Agua (Bubalus Bubalis) ?Documento7 páginas¿Es La Neosporosis Un Problema A Considerar en El Búfalo de Agua (Bubalus Bubalis) ?VenezuelaGanadera.comAún no hay calificaciones
- Genética Molecular para El Mejoramiento de Los Rasgos Reproductivos en BovinosDocumento9 páginasGenética Molecular para El Mejoramiento de Los Rasgos Reproductivos en BovinosVenezuelaGanadera.comAún no hay calificaciones
- Innovadora Técnica de Lactoinducción para Mejorar La Productividad en La Ganadería Doble Propósito MEDICINADocumento9 páginasInnovadora Técnica de Lactoinducción para Mejorar La Productividad en La Ganadería Doble Propósito MEDICINAVenezuelaGanadera.comAún no hay calificaciones
- Resistencia A Enfermedades en La Ganadería Doble Propósito, Un Enfoque GenéticoDocumento10 páginasResistencia A Enfermedades en La Ganadería Doble Propósito, Un Enfoque GenéticoVenezuelaGanadera.comAún no hay calificaciones
- Marcadores Genéticos para El Mejoramiento de La Terneza de La CarneDocumento11 páginasMarcadores Genéticos para El Mejoramiento de La Terneza de La CarneVenezuelaGanadera.comAún no hay calificaciones
- Los Beneficios Del Diagnóstico Molecular en La Ganadería Doble PropósitoDocumento8 páginasLos Beneficios Del Diagnóstico Molecular en La Ganadería Doble PropósitoVenezuelaGanadera.comAún no hay calificaciones
- EndometriosisDocumento37 páginasEndometriosisOsmar RazoAún no hay calificaciones
- Metodo Del Ritmo CalendarioDocumento20 páginasMetodo Del Ritmo CalendarioDaniela LagosAún no hay calificaciones
- UPD Lista de Diálogos SPADocumento6 páginasUPD Lista de Diálogos SPAVictor QueroAún no hay calificaciones
- Sindrome de Ovario PoliquisticoDocumento60 páginasSindrome de Ovario PoliquisticoThabi MonttAún no hay calificaciones
- Rifamicina SV Solución Uso Local Spray PomadaDocumento2 páginasRifamicina SV Solución Uso Local Spray Pomadasiladria100% (1)
- Métodos ANTICONCEPTIVOS - 20231127 - 122151 - 0000Documento1 páginaMétodos ANTICONCEPTIVOS - 20231127 - 122151 - 0000Rosaura PerezAún no hay calificaciones
- 6to Grado - Bloque 1 EXAMENDocumento11 páginas6to Grado - Bloque 1 EXAMENAndyie MorAún no hay calificaciones
- Dispositivos IntrauterinosDocumento42 páginasDispositivos IntrauterinosFernanda AhumadaAún no hay calificaciones
- Rape AxeDocumento9 páginasRape AxeAnonymous CSudLPP1Aún no hay calificaciones
- Condón FemeninoDocumento3 páginasCondón FemeninoSalvador Romero GaliciaAún no hay calificaciones
- Casos Clinicos Alteraciones MenstrualesDocumento31 páginasCasos Clinicos Alteraciones MenstrualesSociedad de Ginecologia de Misiones100% (2)
- Metodos de Planificacion Familiar de BarreraDocumento10 páginasMetodos de Planificacion Familiar de BarreraTahiry Gissel Pax RodriguezAún no hay calificaciones
- Articulo Marketing de CondonesDocumento38 páginasArticulo Marketing de CondonesItzzi CeballosAún no hay calificaciones
- Folleto Metodos AnticonceptivosDocumento2 páginasFolleto Metodos AnticonceptivosCintia SackAún no hay calificaciones
- Triptico de Planificacion FamiliarDocumento4 páginasTriptico de Planificacion FamiliarWENDY MENDOZA100% (1)
- Hemorragia Uterina DisfuncionalDocumento10 páginasHemorragia Uterina DisfuncionalDiana LeidyAún no hay calificaciones
- Ventajas y Desventajas de Los Métodos AnticonceptivosDocumento2 páginasVentajas y Desventajas de Los Métodos AnticonceptivosAnabellaAún no hay calificaciones
- Ciclo Sexual FemeninoDocumento7 páginasCiclo Sexual FemeninoJavier Marcelo AndinoAún no hay calificaciones
- Biología 3° Año Métodos de Control de NatalidadDocumento9 páginasBiología 3° Año Métodos de Control de NatalidadpaulaAún no hay calificaciones
- Monografia Metodos 1Documento44 páginasMonografia Metodos 1padre rumiAún no hay calificaciones
- Falsas Creencias Sobre El Aborto y Su Relaciã N Con La Salud de La Mujer PDFDocumento15 páginasFalsas Creencias Sobre El Aborto y Su Relaciã N Con La Salud de La Mujer PDFANGELAún no hay calificaciones
- Tarjetero de Control de Usuarias y Usuarios de PF ManualDocumento17 páginasTarjetero de Control de Usuarias y Usuarios de PF ManualmarcelaAún no hay calificaciones
- Examen práctica gráficas informaciónDocumento9 páginasExamen práctica gráficas informaciónAna LopezAún no hay calificaciones
- Ciclo Genital FemeninoDocumento19 páginasCiclo Genital FemeninoDeiby TorricoAún no hay calificaciones
- 58-Texto Del Artículo-58-2-10-20200917Documento8 páginas58-Texto Del Artículo-58-2-10-20200917Cristina AstudilloAún no hay calificaciones
- Relaciones Sexuales Entre Parejas HomosexualesDocumento6 páginasRelaciones Sexuales Entre Parejas HomosexualesAlondra ZepedaAún no hay calificaciones