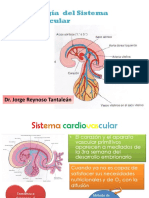
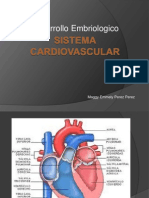
También podría gustarte
- Manual Neuropsi PDFDocumento69 páginasManual Neuropsi PDFmelissa97% (36)
- Sistema CirculatorioDocumento12 páginasSistema CirculatorioJessica Andrea Alamilla GarcíaAún no hay calificaciones
- Experimento RelojDocumento15 páginasExperimento RelojLizz Frizz RockAún no hay calificaciones
- Corazon EmbrionarioDocumento9 páginasCorazon EmbrionarioJhonatan Peña AcasieteAún no hay calificaciones
- Embriología Histologia de CardioDocumento4 páginasEmbriología Histologia de CardiobacagmariafAún no hay calificaciones
- Embriologia Del Sistema CardiovascularDocumento6 páginasEmbriologia Del Sistema CardiovascularNathalia PerezAún no hay calificaciones
- Sistema Cardiovascular. EMBRIOLOGIA PDFDocumento10 páginasSistema Cardiovascular. EMBRIOLOGIA PDFDavid De La RosaAún no hay calificaciones
- Sistema CardiorespiratorioDocumento14 páginasSistema CardiorespiratorioEsteban Ramos MarinAún no hay calificaciones
- Resumen de La Segunda Semana EmbrioDocumento8 páginasResumen de La Segunda Semana EmbrioMario LazoAún no hay calificaciones
- Resumen de TEMA 5 - ORGANOGÉNESISDocumento3 páginasResumen de TEMA 5 - ORGANOGÉNESISAlicia HernandezAún no hay calificaciones
- Clase 11 Guia Grupo 2Documento11 páginasClase 11 Guia Grupo 2María IsabelAún no hay calificaciones
- Embriologia Del Sistema CardiovascularDocumento32 páginasEmbriologia Del Sistema CardiovascularJcaminoAún no hay calificaciones
- Embriología 8Documento51 páginasEmbriología 8Nicole DíazAún no hay calificaciones
- Desarrollo Embriológico Del Sistema CardiovascularDocumento10 páginasDesarrollo Embriológico Del Sistema CardiovascularareloyaezAún no hay calificaciones
- SISTEMA CARDIOVASCULAR UwUDocumento13 páginasSISTEMA CARDIOVASCULAR UwUPaola AvilaAún no hay calificaciones
- Embriología: Sistema Cardiovascular PresentaciónDocumento11 páginasEmbriología: Sistema Cardiovascular PresentaciónEverardo MartinezAún no hay calificaciones
- CorazonDocumento14 páginasCorazonYareli PerezAún no hay calificaciones
- Sistema Cardiovascular-EmbriologiaDocumento185 páginasSistema Cardiovascular-EmbriologiaLiliana Anglas SilvaAún no hay calificaciones
- Sesion 4 Tercerasemanade Desarrollo EmbrionarioDocumento16 páginasSesion 4 Tercerasemanade Desarrollo EmbrionarioAndrea Nickole MedinaAún no hay calificaciones
- Desarrollo Corazon 1 OdoDocumento19 páginasDesarrollo Corazon 1 OdoAlexander Ivan Cornejo SanchezAún no hay calificaciones
- Sistema Cardiovascular EmbriologíaDocumento4 páginasSistema Cardiovascular EmbriologíaLuna JoyaAún no hay calificaciones
- Embriología CorazónDocumento15 páginasEmbriología CorazónPedro PlataAún no hay calificaciones
- Trabajo de Investigación Fisiología MédicaDocumento8 páginasTrabajo de Investigación Fisiología MédicaDIEGO OSWALDO MAYAGOITIA TORTOLEDOAún no hay calificaciones
- Desarrollo Del Sistema CardiovascularDocumento11 páginasDesarrollo Del Sistema CardiovascularAlicia Hernández PérezAún no hay calificaciones
- Anatomía Del Desarrollo 2Documento45 páginasAnatomía Del Desarrollo 2nimAún no hay calificaciones
- Aparato Cardiovascular 2 Embrio 2023Documento6 páginasAparato Cardiovascular 2 Embrio 2023RaoulAún no hay calificaciones
- UABP 2. Aparato CardiovascularDocumento6 páginasUABP 2. Aparato CardiovascularMartín PanicoAún no hay calificaciones
- Corazón: Tubos EndocárdicosDocumento3 páginasCorazón: Tubos Endocárdicosromimi fAún no hay calificaciones
- Sistema CardiovascularDocumento6 páginasSistema Cardiovascularvanessa cruz gomezAún no hay calificaciones
- Desarrollo Del Sistema CardiovascularDocumento3 páginasDesarrollo Del Sistema CardiovascularFrancisco MoroyoquiAún no hay calificaciones
- Resumen de Embriologia 2do ParcialDocumento36 páginasResumen de Embriologia 2do ParcialMigela Belen Rojas DelgadoAún no hay calificaciones
- Cardiovascular (EM Carlos y HibDocumento40 páginasCardiovascular (EM Carlos y HibGabi FerreroAún no hay calificaciones
- Cardio Embrio ClaseDocumento68 páginasCardio Embrio Claseluis fernandoAún no hay calificaciones
- Resumen Capitulo 17 Langman - Formación Del Sistema CardiacoDocumento5 páginasResumen Capitulo 17 Langman - Formación Del Sistema Cardiacorafael mendezAún no hay calificaciones
- Desarrollo de Los SomitasDocumento3 páginasDesarrollo de Los SomitasPedro Murillo GAún no hay calificaciones
- Sistema CardiovascularDocumento7 páginasSistema CardiovascularJoseph FaríasAún no hay calificaciones
- Sistema Cardiovascular - Embriologã - ADocumento52 páginasSistema Cardiovascular - Embriologã - Acarlos ramon de sousa gomesAún no hay calificaciones
- Guía de Práctica 12 2023Documento13 páginasGuía de Práctica 12 2023JAROLL JHANPOLL CUZCO MENDOZAAún no hay calificaciones
- Sistema Embriologico CardioRespiratorioDocumento44 páginasSistema Embriologico CardioRespiratorioleonelknightAún no hay calificaciones
- Resumen Sistema CardiovascularDocumento14 páginasResumen Sistema CardiovascularVanessa CastañedaAún no hay calificaciones
- Resumen de Cardiologia-1Documento5 páginasResumen de Cardiologia-1jennifer garciaAún no hay calificaciones
- Sistema Arterial EmbrioDocumento51 páginasSistema Arterial EmbrioMariano J. CalderónAún no hay calificaciones
- Embriologia Del Sistema CardiovascularDocumento13 páginasEmbriologia Del Sistema CardiovascularMirgelys VargasAún no hay calificaciones
- Desarrollo Embriológico Del Corazón y PericardioDocumento6 páginasDesarrollo Embriológico Del Corazón y PericardioYlianaAún no hay calificaciones
- Sistema CardiovascularDocumento12 páginasSistema CardiovascularCristina VelázquezAún no hay calificaciones
- Embriologia Sistema CardiovascularDocumento21 páginasEmbriologia Sistema CardiovascularAngie BurbanoAún no hay calificaciones
- Embriologia Del Sistema CardiovascularDocumento17 páginasEmbriologia Del Sistema CardiovascularTomas GomezAún no hay calificaciones
- Taller de Embriología CardiorespiratoriaDocumento16 páginasTaller de Embriología Cardiorespiratorialeiny camila bAún no hay calificaciones
- Sistema CardiovascularDocumento11 páginasSistema CardiovascularDaav L. GuarneriAún no hay calificaciones
- 07-UNIDAD III Embriologia Del CorazonDocumento56 páginas07-UNIDAD III Embriologia Del CorazonCesia J. Darce100% (1)
- Embriogénesis CardiovascularDocumento4 páginasEmbriogénesis CardiovascularJulieta MoraAún no hay calificaciones
- Desarrollo Embrionario en La Tercera SemanaDocumento28 páginasDesarrollo Embrionario en La Tercera SemanaCristellE.AguilarManjarrez0% (1)
- Sistema CardiovascularDocumento29 páginasSistema CardiovascularJaime Bodipo NzambiAún no hay calificaciones
- Actividad 1. Embriologia CardiovascularDocumento10 páginasActividad 1. Embriologia Cardiovascularkarime marquezAún no hay calificaciones
- Sistema CardiovascularDocumento20 páginasSistema CardiovascularmaggyemmelyAún no hay calificaciones
- Embriología Clase 2Documento19 páginasEmbriología Clase 2MARIANA ESTRADA RESTREPOAún no hay calificaciones
- Embriología Sistema CardiovascularDocumento3 páginasEmbriología Sistema Cardiovascularalejandra rubensAún no hay calificaciones
- Clase Viii Embriologia CardiovascularDocumento62 páginasClase Viii Embriologia CardiovascularAnahi CaizaAún no hay calificaciones
- Cavidades CorporalesDocumento50 páginasCavidades CorporalesCarlos Antonio Sauco CastroAún no hay calificaciones
- Anatomía Del CorazónDocumento5 páginasAnatomía Del CorazónVeronica perezAún no hay calificaciones
- Guia Parcial 2 EmbrioDocumento4 páginasGuia Parcial 2 EmbrioDagoberto GarzaAún no hay calificaciones
- Memorial GobernacionDocumento7 páginasMemorial GobernacionGustavo BallivianAún no hay calificaciones
- Diagrama V de Gowin Plantilla AlumnosDocumento1 páginaDiagrama V de Gowin Plantilla Alumnosrobe535100% (1)
- Joven VecinoDocumento2 páginasJoven VecinoAlbert Daker Ticona HuaytaAún no hay calificaciones
- Test DesiderativoDocumento6 páginasTest DesiderativoChristian Algarañaz FernandezAún no hay calificaciones
- Corrosión de Los Envases de HojalataDocumento22 páginasCorrosión de Los Envases de HojalataBrayan A. Damian TolentinoAún no hay calificaciones
- Ficha Técnica Caneca Plásica 55 Galones 01 TAI 208 11 Ene 20Documento1 páginaFicha Técnica Caneca Plásica 55 Galones 01 TAI 208 11 Ene 20Julio BuendiaAún no hay calificaciones
- Reproduccion de Plantas y AnimalesDocumento19 páginasReproduccion de Plantas y AnimalesCeledonio CamanaAún no hay calificaciones
- Ensayos en Cables de Media Tensión.: Centelsa Xlpe Cu 90 °C 250 Kcmil 15 KV 100% PC1 HDPEDocumento33 páginasEnsayos en Cables de Media Tensión.: Centelsa Xlpe Cu 90 °C 250 Kcmil 15 KV 100% PC1 HDPEomar100% (2)
- Etiquetado de FertilizantesDocumento4 páginasEtiquetado de FertilizantesRafael Mejia AmayaAún no hay calificaciones
- PSST-02 Comunicación Externa e InternaDocumento4 páginasPSST-02 Comunicación Externa e InternaJonathan Alexander Moreno MartinezAún no hay calificaciones
- Maria Montessori, Biografia.Documento2 páginasMaria Montessori, Biografia.Fernanda EscalonaAún no hay calificaciones
- Conocimiento Dodontolgico en Niños Con SD-cirujanos DentistasDocumento79 páginasConocimiento Dodontolgico en Niños Con SD-cirujanos Dentistashector rodriguez neyraAún no hay calificaciones
- AMILASADocumento1 páginaAMILASAWendy MaciasAún no hay calificaciones
- Proyecto de Aula Entrega FinalDocumento26 páginasProyecto de Aula Entrega FinalSergio Alejandro Severiche GuerreroAún no hay calificaciones
- 2021 AplazadosDocumento2 páginas2021 AplazadosRenzo Jair Arestegui TacuriAún no hay calificaciones
- Ley Que Modifica Artículo 11 de La Ley General de SaludDocumento3 páginasLey Que Modifica Artículo 11 de La Ley General de SaludVary H. Silva SerrotAún no hay calificaciones
- Lectura 12 ADULTEZ TEMPRANA PDFDocumento10 páginasLectura 12 ADULTEZ TEMPRANA PDFRoxana AscaAún no hay calificaciones
- M10 U1 S2 FelbDocumento15 páginasM10 U1 S2 FelbFher LopezAún no hay calificaciones
- Cálculo de Dispersión HCN para Liberaciones Instantaneas (Puff)Documento9 páginasCálculo de Dispersión HCN para Liberaciones Instantaneas (Puff)Jhon Jacome Giraldo100% (1)
- Guia 20Documento6 páginasGuia 20YosaharaVidalMateoAún no hay calificaciones
- Caso Clinico - Insuficiencia Renal AgudaDocumento5 páginasCaso Clinico - Insuficiencia Renal AgudaNAZARETH DAHANA ORTIZ NUÑEZAún no hay calificaciones
- Trabajo Practico - Teorias de La PenaDocumento22 páginasTrabajo Practico - Teorias de La PenaSantiago Juarros100% (1)
- Exploración Clínica Del Globo OcularDocumento7 páginasExploración Clínica Del Globo OcularAlexis TkmAún no hay calificaciones
- 3 - Quimica General GasesDocumento11 páginas3 - Quimica General Gasesmajo silvaAún no hay calificaciones
- Salud Pública - Avanzado - Sim Especialidad Rm21 - Sin ClavesDocumento8 páginasSalud Pública - Avanzado - Sim Especialidad Rm21 - Sin ClavesfransforAún no hay calificaciones
- Acta de Buena ConductaDocumento10 páginasActa de Buena Conductavictor mamani100% (1)
- Power Point Tesis Unerg Presentacion Final (Formato 2010) - 3 1Documento20 páginasPower Point Tesis Unerg Presentacion Final (Formato 2010) - 3 1Tibisay AvendañoAún no hay calificaciones
- Mdae15t5caso Grupo 108Documento7 páginasMdae15t5caso Grupo 108luisAún no hay calificaciones