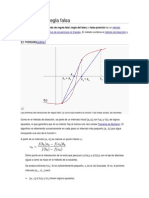
También podría gustarte
- Modelos Cinetica EnzimaticaDocumento4 páginasModelos Cinetica EnzimaticaDui Dui Dui DuiAún no hay calificaciones
- Tema 012Documento27 páginasTema 012Franz Tucta HuillcaAún no hay calificaciones
- Tema 012Documento27 páginasTema 012Luis Salinas AAún no hay calificaciones
- Bioquimica WordDocumento3 páginasBioquimica WordDaniela Valencia LópezAún no hay calificaciones
- Determinación de Km y Vmax mediante métodos gráficos enzimáticosDocumento2 páginasDeterminación de Km y Vmax mediante métodos gráficos enzimáticosEdson DiazAún no hay calificaciones
- Cinética de Las Reacciones para Sistemas BiológicosDocumento41 páginasCinética de Las Reacciones para Sistemas BiológicosJessy HernándezAún no hay calificaciones
- Ingenieria de Reacciones Enzimàticas: Desarrollo de Expresiones Cinèticas.Documento21 páginasIngenieria de Reacciones Enzimàticas: Desarrollo de Expresiones Cinèticas.Balam ToscanoAún no hay calificaciones
- Práctica Métodos Experimentales de Parámetros CinéticosDocumento4 páginasPráctica Métodos Experimentales de Parámetros CinéticosMarianAún no hay calificaciones
- Método de La Regla FalsaDocumento2 páginasMétodo de La Regla FalsaSergioGomezAún no hay calificaciones
- Método de La Regla FalsaDocumento2 páginasMétodo de La Regla FalsaJavier Salvador Martinez SalasAún no hay calificaciones
- EDocumento1 páginaEGuadalupe GAún no hay calificaciones
- El Método Más Simple e Intuitivo de Integración Aproximada Es El Método de Los TrapeciosDocumento5 páginasEl Método Más Simple e Intuitivo de Integración Aproximada Es El Método de Los TrapeciosNataly BayasAún no hay calificaciones
- Determinación de la cinética enzimática de la invertasa mediante el análisis de Michaelis-MentenDocumento6 páginasDeterminación de la cinética enzimática de la invertasa mediante el análisis de Michaelis-MentenDago OrtizAún no hay calificaciones
- Determinación de La Concentración de Diluciones de Permanganato de Potasio-1Documento3 páginasDeterminación de La Concentración de Diluciones de Permanganato de Potasio-1Juan RenAún no hay calificaciones
- Demostración de Ecuaciones de Pendiente DeflexiónDocumento12 páginasDemostración de Ecuaciones de Pendiente DeflexiónJoseluish ItapizacoAún no hay calificaciones
- Ec MichaelisDocumento10 páginasEc MichaelisJeka JekatronikAún no hay calificaciones
- Ecuación Van Der Waals resuelta numéricamenteDocumento10 páginasEcuación Van Der Waals resuelta numéricamenteCitlali Jazmín100% (1)
- Relación lineal entre distancia y tiempo de una burbuja de aireDocumento8 páginasRelación lineal entre distancia y tiempo de una burbuja de aireMitzi Moreno0% (1)
- Ley de Ohm1Documento2 páginasLey de Ohm1musa saiehAún no hay calificaciones
- Informe 9Documento14 páginasInforme 9Gaby CampoverdeAún no hay calificaciones
- Calculo de Vigas ContinuasDocumento9 páginasCalculo de Vigas ContinuasYesica Tatiana100% (4)
- Método de Volúmenes FinitosDocumento8 páginasMétodo de Volúmenes FinitosFelipe Carlo Galdames OlmedoAún no hay calificaciones
- ENZIMAS - Biotecnología 2.1Documento78 páginasENZIMAS - Biotecnología 2.1BarbyModestiAún no hay calificaciones
- Campo de AceleraciónDocumento6 páginasCampo de AceleraciónLuis MuñozAún no hay calificaciones
- Operaciones Con Ondas Ing ElectricaDocumento6 páginasOperaciones Con Ondas Ing ElectricaRudy Q.VargasAún no hay calificaciones
- CINÉTICA ENZIMÁTICA - Biologia y QuimicaDocumento12 páginasCINÉTICA ENZIMÁTICA - Biologia y QuimicaWILDER PIZARRO CUNYASAún no hay calificaciones
- Actividad Enzimática de La Fosfatasa AlcalinaDocumento16 páginasActividad Enzimática de La Fosfatasa AlcalinaDayana Florez GamboaAún no hay calificaciones
- Metodo Diferencial de Analisis de DatosDocumento17 páginasMetodo Diferencial de Analisis de DatosRocio Perez Callejas100% (1)
- Determinaciòn de Los Parámetros Cinéticos de La EnzimaDocumento30 páginasDeterminaciòn de Los Parámetros Cinéticos de La EnzimaLeon VásquezAún no hay calificaciones
- Separación de La Ecuación Independiente Del TiempoDocumento10 páginasSeparación de La Ecuación Independiente Del TiempoDianny 97Aún no hay calificaciones
- 4) El Metodo Vectorial y El Metodo Sibolico 2Documento4 páginas4) El Metodo Vectorial y El Metodo Sibolico 2'LautaroMedranoAún no hay calificaciones
- Construcción de Gráficas y Ecuaciones EmpíricasDocumento23 páginasConstrucción de Gráficas y Ecuaciones EmpíricasAlexander Chavez EspinozaAún no hay calificaciones
- Proyecto Fenomenos de Transporte IDocumento2 páginasProyecto Fenomenos de Transporte IDUBAN ANDRES CERON MARTINEZAún no hay calificaciones
- Resumenes y ProblemasDocumento16 páginasResumenes y ProblemasJavier RodriguezAún no hay calificaciones
- Forma Integrada de La Ecuación de MichaelisDocumento5 páginasForma Integrada de La Ecuación de MichaelisAngela CaceresAún no hay calificaciones
- Trabajo 01 Analisis 1Documento13 páginasTrabajo 01 Analisis 1ROSMERY BOZA BENDEZUAún no hay calificaciones
- Ecuaciones de Michaelis PDFDocumento2 páginasEcuaciones de Michaelis PDFAmaury Daniel CortesAún no hay calificaciones
- Reacciones MultisustratoDocumento12 páginasReacciones MultisustratoKarla Alejo GonzalezAún no hay calificaciones
- CFD Comparación Modelos Upwind, QUICK, y Centradas Por Volumenes FinitosDocumento15 páginasCFD Comparación Modelos Upwind, QUICK, y Centradas Por Volumenes FinitosJohn Jairo Ortiz MartinezAún no hay calificaciones
- Tuberias Ramificadas Problema 3 TanquesDocumento42 páginasTuberias Ramificadas Problema 3 TanquesJuan Carlos Pedroza GarridoAún no hay calificaciones
- Cinética Enzimática Trabajo CorregidoDocumento16 páginasCinética Enzimática Trabajo CorregidoTATIANA MARCELA BERMEJO ARAUJOAún no hay calificaciones
- Copia de Cinetica EnzimáticaDocumento6 páginasCopia de Cinetica EnzimáticaYohan MoralesAún no hay calificaciones
- Métodos de LinealizaciónDocumento5 páginasMétodos de LinealizaciónDanny Paul100% (2)
- HLKHKDocumento17 páginasHLKHKLuc VilcaAún no hay calificaciones
- Modelos para La Conpensacion de AngulosDocumento4 páginasModelos para La Conpensacion de AngulosJesus Simeon de WhitteAún no hay calificaciones
- Método de Los Tres MomentosDocumento6 páginasMétodo de Los Tres Momentosjhon ivanAún no hay calificaciones
- Determinación de los parámetros Vmax y Km de la ecuación de Michaelis-MentenDocumento5 páginasDeterminación de los parámetros Vmax y Km de la ecuación de Michaelis-MentenAndrés Felipe Aristizabal MonterrozaAún no hay calificaciones
- Mecapedia-Cálculo de La Catenaria Conocidos Los Puntos de Amarre y La LongitudDocumento4 páginasMecapedia-Cálculo de La Catenaria Conocidos Los Puntos de Amarre y La LongitudErnesto Simón Ascencios Zorrilla100% (1)
- Cuestionario enzimas: energía de activación, equilibrio químico y cinética enzimáticaDocumento5 páginasCuestionario enzimas: energía de activación, equilibrio químico y cinética enzimáticaNestor RuizAún no hay calificaciones
- UntitledDocumento10 páginasUntitledÁngel MartínezAún no hay calificaciones
- Respuestas Jouglard Actualizado 19-12-13 CalculoDocumento4 páginasRespuestas Jouglard Actualizado 19-12-13 CalculoSantiSilveyraAún no hay calificaciones
- Componentes SimetricasDocumento14 páginasComponentes SimetricasCRISTIANAún no hay calificaciones
- Metodos NumericosDocumento8 páginasMetodos NumericosLeonardo RdzAún no hay calificaciones
- Ejercicios de Vectores, Mru, MruaDocumento13 páginasEjercicios de Vectores, Mru, MruaBayron Leandro Rosero Huertas100% (1)
- I. Seminarios Cinética EnzimáticaDocumento7 páginasI. Seminarios Cinética EnzimáticaLaura Célix100% (1)
- Regresión lineal aplicada a la ingeniería químicaDocumento24 páginasRegresión lineal aplicada a la ingeniería químicaJosé Arteaga0% (1)
- CampoVelocidadFluidosMatlabDocumento7 páginasCampoVelocidadFluidosMatlabLuis MuñozAún no hay calificaciones
- Análisis de fuerzas en dos y tres dimensionesDocumento87 páginasAnálisis de fuerzas en dos y tres dimensionesIRLANDAAún no hay calificaciones
- Trafico Intracelular de ProteinasDocumento27 páginasTrafico Intracelular de ProteinasPhillip David Ormeño Vasquez0% (1)
- TrascripcionDocumento27 páginasTrascripcionyatoAún no hay calificaciones
- Producción PHA Bacillus glicerolDocumento9 páginasProducción PHA Bacillus glicerolAngela GarciaAún no hay calificaciones
- Word Desplazamiento Miscible e InmiscibleDocumento19 páginasWord Desplazamiento Miscible e InmiscibleRoronoazoro67% (3)
- Grupos FuncionalesDocumento2 páginasGrupos FuncionalesDaniela SantisbonAún no hay calificaciones
- Aines en Odontologia IMPORTANTEDocumento34 páginasAines en Odontologia IMPORTANTEOrlando RodríguezAún no hay calificaciones
- Biopolimeros InformeDocumento58 páginasBiopolimeros InformePATRICIA ROSARIO SANTANDER SALASAún no hay calificaciones
- TRABAJO PRÁCTICO Introducción A La QuímicaDocumento5 páginasTRABAJO PRÁCTICO Introducción A La QuímicamariaAún no hay calificaciones
- Aprovechamiento de ResiduosDocumento129 páginasAprovechamiento de Residuosandrea0% (1)
- Unidad II Anatomia Y Fisiología de La CelulaDocumento26 páginasUnidad II Anatomia Y Fisiología de La Celulaana yuivi yucraAún no hay calificaciones
- Informe Reacción de HenryDocumento10 páginasInforme Reacción de HenryJhon GalloAún no hay calificaciones
- Nociones Fundamentales de La Quimica BioDocumento321 páginasNociones Fundamentales de La Quimica BioKatie MhAún no hay calificaciones
- Clasificacion Del PetroleoDocumento11 páginasClasificacion Del PetroleoRox LopezAún no hay calificaciones
- Universidad Central Del Ecuador: Características MorfológicasDocumento4 páginasUniversidad Central Del Ecuador: Características MorfológicasAlex OwenAún no hay calificaciones
- Mosaico FluidoDocumento24 páginasMosaico Fluidociber konektAún no hay calificaciones
- PRÁCTICA 8 Separacion de ProteinasDocumento1 páginaPRÁCTICA 8 Separacion de Proteinasventaquemada boyacaAún no hay calificaciones
- Descripción 4 PDFDocumento116 páginasDescripción 4 PDFDavid Aristizabal VelásquezAún no hay calificaciones
- Cuestiones tipo test PAEG sobre agua, sales, glúcidos y lípidos (menos deDocumento66 páginasCuestiones tipo test PAEG sobre agua, sales, glúcidos y lípidos (menos deJackelineAún no hay calificaciones
- Producción de acetato de sodio en CSTRDocumento5 páginasProducción de acetato de sodio en CSTROscar ToroAún no hay calificaciones
- Análisis cenizas alimentosDocumento7 páginasAnálisis cenizas alimentosPerlera AnnyAún no hay calificaciones
- 1practica 1 BioquimicaDocumento9 páginas1practica 1 BioquimicaXIMENAAún no hay calificaciones
- Reacciones de adición 1QM2Documento5 páginasReacciones de adición 1QM2Odalhyz CabreraAún no hay calificaciones
- Motores Diesel...Documento22 páginasMotores Diesel...wrallan0% (1)
- 2.2.9 Higiene y Seguridad NormasDocumento7 páginas2.2.9 Higiene y Seguridad NormasCarlos OlmecaAún no hay calificaciones
- Guia de Resultados Practica 5 AromaticosDocumento5 páginasGuia de Resultados Practica 5 AromaticosCarlos AmadorAún no hay calificaciones
- Biologia Selectividad Examen 7 Resuelto Castilla y Leon WWW - Siglo21x.blogspotDocumento7 páginasBiologia Selectividad Examen 7 Resuelto Castilla y Leon WWW - Siglo21x.blogspotmonografiasxAún no hay calificaciones
- Carbohidratos ZootecniaDocumento2 páginasCarbohidratos ZootecniaJohan MorenoAún no hay calificaciones
- Vitaminas y enfermedades por deficienciaDocumento2 páginasVitaminas y enfermedades por deficienciaMajitito Taipe A100% (1)
- Cuestionario Compuestos AromaticosDocumento6 páginasCuestionario Compuestos AromaticosSebastian torruco gonzalez100% (1)
- Nitración de Benceno Lab.22Documento11 páginasNitración de Benceno Lab.22Diego Medrano Hidalgo100% (1)
- Disciplina Mental: Técnicas infalibles para lograr todo lo que te propones y eliminar la pereza y la procrastinación de tu vida para siempreDe EverandDisciplina Mental: Técnicas infalibles para lograr todo lo que te propones y eliminar la pereza y la procrastinación de tu vida para siempreCalificación: 5 de 5 estrellas5/5 (3)
- La revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaDe EverandLa revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaCalificación: 5 de 5 estrellas5/5 (200)
- La metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceDe EverandLa metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceCalificación: 5 de 5 estrellas5/5 (8)
- Reconstrucción de dientes endodonciados: Pautas de actuación clínicaDe EverandReconstrucción de dientes endodonciados: Pautas de actuación clínicaCalificación: 5 de 5 estrellas5/5 (4)
- Zensorialmente : Dejá que tu cuerpo sea tu cerebroDe EverandZensorialmente : Dejá que tu cuerpo sea tu cerebroAún no hay calificaciones
- Anatomía & 100 estiramientos Esenciales (Color): Técnicas, beneficios, precauciones, consejos, tablas de series, dolenciasDe EverandAnatomía & 100 estiramientos Esenciales (Color): Técnicas, beneficios, precauciones, consejos, tablas de series, dolenciasCalificación: 4.5 de 5 estrellas4.5/5 (21)
- Fisiopatología de las enfermedades cardiovascularesDe EverandFisiopatología de las enfermedades cardiovascularesCalificación: 5 de 5 estrellas5/5 (1)
- El libro conciso de la punción seca: Manual del terapeuta para las aplicaciones en los puntos gatillo miofasciales (Color)De EverandEl libro conciso de la punción seca: Manual del terapeuta para las aplicaciones en los puntos gatillo miofasciales (Color)Calificación: 3 de 5 estrellas3/5 (2)
- Interpretación del ECG: Una Guía Práctica e Intuitiva para Aprender a Leer el ECG y Diagnosticar y Tratar ArritmiasDe EverandInterpretación del ECG: Una Guía Práctica e Intuitiva para Aprender a Leer el ECG y Diagnosticar y Tratar ArritmiasAún no hay calificaciones
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- Desastres y emergencias. Prevención, mitigación y preparaciónDe EverandDesastres y emergencias. Prevención, mitigación y preparaciónCalificación: 4 de 5 estrellas4/5 (5)
- Prescripción de ejercico físico para la saludDe EverandPrescripción de ejercico físico para la saludCalificación: 5 de 5 estrellas5/5 (1)
- Trucos para Leer la Mente de los Demás: Cómo Adivinar el Pensamiento de los Demás con Poco Esfuerzo. 2 Libros en 1 - Secretos de la Psicología Oscura, Cómo ser un Detector de MentirasDe EverandTrucos para Leer la Mente de los Demás: Cómo Adivinar el Pensamiento de los Demás con Poco Esfuerzo. 2 Libros en 1 - Secretos de la Psicología Oscura, Cómo ser un Detector de MentirasCalificación: 4.5 de 5 estrellas4.5/5 (3)
- Dieta Para El Reflujo Biliar y Gastritis Alcalina - Incluye 20 Deliciosas Recetas Libres de Gluten y de Lácteos Para Tratar y Aliviar el Reflujo Biliar y Sus Molestos SíntomasDe EverandDieta Para El Reflujo Biliar y Gastritis Alcalina - Incluye 20 Deliciosas Recetas Libres de Gluten y de Lácteos Para Tratar y Aliviar el Reflujo Biliar y Sus Molestos SíntomasCalificación: 4 de 5 estrellas4/5 (9)
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- Neurociencia para vencer la depresión: La esprial ascendenteDe EverandNeurociencia para vencer la depresión: La esprial ascendenteCalificación: 4.5 de 5 estrellas4.5/5 (10)
- La vibración de las ondas cerebrales: Recuperar el ritmo de una vida saludable y felizDe EverandLa vibración de las ondas cerebrales: Recuperar el ritmo de una vida saludable y felizCalificación: 5 de 5 estrellas5/5 (7)
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)De EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Calificación: 5 de 5 estrellas5/5 (9)
- Notas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesDe EverandNotas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesAún no hay calificaciones
- El péndulo de sanación: Péndulo hebreo. Investigación y sistematización de la técnicaDe EverandEl péndulo de sanación: Péndulo hebreo. Investigación y sistematización de la técnicaCalificación: 4.5 de 5 estrellas4.5/5 (27)