También podría gustarte
- Bioquímica GenéticaDocumento57 páginasBioquímica GenéticaMaria Paz MercadoAún no hay calificaciones
- 1era Clase de Biología CelularDocumento76 páginas1era Clase de Biología CelularMaria Belen MartinezAún no hay calificaciones
- Exposición Nutrigenética y NutrigenómicaDocumento18 páginasExposición Nutrigenética y NutrigenómicaDiego SteffalooAún no hay calificaciones
- Codigo Genetico Dra. Laura PDFDocumento64 páginasCodigo Genetico Dra. Laura PDFEMELY YAHAYRA TIRADO DIAZAún no hay calificaciones
- Semana 7 - Teoria PB1 WA PDFDocumento40 páginasSemana 7 - Teoria PB1 WA PDFAEI OUAún no hay calificaciones
- Clase 23. ADN. Cromosomas y Cromatina.Documento28 páginasClase 23. ADN. Cromosomas y Cromatina.MelodyAún no hay calificaciones
- Genetica MicrobianaDocumento16 páginasGenetica MicrobianaKevin Flores88% (8)
- Estructura Molecular de Genes y Cormosomas, Cariotipos y PatogeniasDocumento28 páginasEstructura Molecular de Genes y Cormosomas, Cariotipos y PatogeniasChristian SDAún no hay calificaciones
- Manual GenéticaDocumento110 páginasManual GenéticaSergio Alejandro VillarAún no hay calificaciones
- Naturaleza de Gen y GenomaDocumento31 páginasNaturaleza de Gen y GenomaMaria Fernanda Pozo AguirreAún no hay calificaciones
- U6 GeneticaMicrobiana 19166 PDFDocumento85 páginasU6 GeneticaMicrobiana 19166 PDFTottoObandoAún no hay calificaciones
- Semana 14Documento27 páginasSemana 14Mariangeles Loayza MamaniAún no hay calificaciones
- Acidos Nucléicos 20-12-18Documento88 páginasAcidos Nucléicos 20-12-18Yessenia FloresAún no hay calificaciones
- Clase 1 Dna Como Material GeneticoDocumento85 páginasClase 1 Dna Como Material GeneticoNAYELIAún no hay calificaciones
- TranscripcionDocumento29 páginasTranscripcionalicelopes0905Aún no hay calificaciones
- Unidad 3. BH 2019Documento42 páginasUnidad 3. BH 2019More AguirreAún no hay calificaciones
- Taller 3 FinalDocumento55 páginasTaller 3 FinalAlejandro García RigualAún no hay calificaciones
- Bases de La Genética MicrobianaDocumento25 páginasBases de La Genética MicrobianaVALERIA GRAún no hay calificaciones
- Clase 8 y 9 - Replicación, Transcripción y TraducciónDocumento24 páginasClase 8 y 9 - Replicación, Transcripción y TraducciónAndrés Sepúlveda NavarroAún no hay calificaciones
- Genetica Bacteriana. TextoDocumento46 páginasGenetica Bacteriana. TextoMagdalena Oñate SerranoAún no hay calificaciones
- Genética Bacteriana I 2016Documento53 páginasGenética Bacteriana I 2016Valeria Dos SantosAún no hay calificaciones
- Neticamicrobiana 21174 PDFDocumento84 páginasNeticamicrobiana 21174 PDFVanessa SánchezAún no hay calificaciones
- Del ADN Al Fenotipo - eDocumento35 páginasDel ADN Al Fenotipo - eDavid Esteban Cantor AponteAún no hay calificaciones
- Repaso 1, GenéticaDocumento7 páginasRepaso 1, GenéticaBRANDON YAHIR CADENA LOPEZAún no hay calificaciones
- Gene Genomas ADN y ReplicacionDocumento52 páginasGene Genomas ADN y ReplicacionMishel Jordan ZeballosAún no hay calificaciones
- Organización Del GenomaDocumento40 páginasOrganización Del GenomaMariana PuertasAún no hay calificaciones
- Clase 2 de MicrobiologíaDocumento97 páginasClase 2 de MicrobiologíaRusvel Alex Vilca SivirichiAún no hay calificaciones
- GENÉTICADocumento5 páginasGENÉTICANaivy PulidoAún no hay calificaciones
- Clase 13 Sin AudioDocumento66 páginasClase 13 Sin Audiomillaray.medinaAún no hay calificaciones
- Genes IDocumento45 páginasGenes IGian LuccaAún no hay calificaciones
- Guia BiocelDocumento11 páginasGuia BiocelEstefania Alexandra Del Villar RodríguezAún no hay calificaciones
- Organización de Los Genomas 2019Documento63 páginasOrganización de Los Genomas 2019Laura100% (1)
- Intro Genetica 2022Documento46 páginasIntro Genetica 2022Ivan MolinaAún no hay calificaciones
- A-02 Conceptos Basicos Transcripciã NDocumento16 páginasA-02 Conceptos Basicos Transcripciã NOSMAR ZARATE LOPEZAún no hay calificaciones
- Genética Molecular I. Replicación y Transcripción Del Adn.Documento10 páginasGenética Molecular I. Replicación y Transcripción Del Adn.Claudia Prieto MontesAún no hay calificaciones
- Material 2019F1 BIO101 01 129641Documento35 páginasMaterial 2019F1 BIO101 01 129641Ciudadana CelestialAún no hay calificaciones
- ADN y Codigo Genetico 120929Documento74 páginasADN y Codigo Genetico 120929Astrid CárdenasAún no hay calificaciones
- Resumen Hibridación y PCRDocumento14 páginasResumen Hibridación y PCRkarlotamgAún no hay calificaciones
- Genetica BacterianaDocumento20 páginasGenetica Bacterianaflorencia bertoldiAún no hay calificaciones
- Ensayo Sobre Gen y GenomaDocumento3 páginasEnsayo Sobre Gen y GenomaDIEGO JAIME THOMASAún no hay calificaciones
- 1 SG1 2013.ppt - 0Documento38 páginas1 SG1 2013.ppt - 0Edu SancariAún no hay calificaciones
- GenéticaDocumento133 páginasGenéticaalexandraAún no hay calificaciones
- Conceptos Basicos MolecularDocumento28 páginasConceptos Basicos Molecularsnoggle.yumiAún no hay calificaciones
- Clase 5 Del ADN A Las ProteinasDocumento53 páginasClase 5 Del ADN A Las ProteinasBarbara BravoAún no hay calificaciones
- El Camino Del ADN A Las ProteínasDocumento17 páginasEl Camino Del ADN A Las ProteínasValentina CicaniaAún no hay calificaciones
- Genetica MicrobianaDocumento71 páginasGenetica MicrobianaKNayAún no hay calificaciones
- 3 Control Genético CelularDocumento49 páginas3 Control Genético CelularEduardo Segura MoralesAún no hay calificaciones
- Semana3 Sesión5 TEO Transcripción, Código Genético y TraducciónDocumento120 páginasSemana3 Sesión5 TEO Transcripción, Código Genético y TraducciónDave DiosesAún no hay calificaciones
- Genetica MolecularDocumento23 páginasGenetica MolecularFlakito Alvarado OsorioAún no hay calificaciones
- Acidos Nucleicos A 2023Documento44 páginasAcidos Nucleicos A 2023Eloina HerreraAún no hay calificaciones
- 13-02-5 BiDocumento96 páginas13-02-5 BicarolinaAún no hay calificaciones
- 4 º Ud.8.Inf - GeneticaDocumento84 páginas4 º Ud.8.Inf - GeneticaMario Deusto RíosAún no hay calificaciones
- Regulación de La ReplicaciónDocumento30 páginasRegulación de La ReplicaciónSpartan CarmineAún no hay calificaciones
- Tema2.IntroducciónGenómica GenómicaEstructural PDFDocumento101 páginasTema2.IntroducciónGenómica GenómicaEstructural PDFJose Antonio Vargas GonzalezAún no hay calificaciones
- Genética Bioquímica II DEFINITIVODocumento195 páginasGenética Bioquímica II DEFINITIVOapi-26262344100% (4)
- Expo Genetica Herencia Extranuclear 15julDocumento14 páginasExpo Genetica Herencia Extranuclear 15julFaviola BertrezAún no hay calificaciones
- Clase 4 TranscripcionDocumento21 páginasClase 4 TranscripcionAzul ParraAún no hay calificaciones
- 1.1 Dogma Central de La Biología Molecular AlumnosDocumento45 páginas1.1 Dogma Central de La Biología Molecular AlumnosSergio CastellanosAún no hay calificaciones
- Tema 1 Biologia Genetica FarmaciaDocumento4 páginasTema 1 Biologia Genetica FarmaciaRaquelAún no hay calificaciones
- Más allá del ADN: La Revolución Epigenética: Desde Mecanismos Celulares hasta Factores Ambientales: Cómo la Epigenética Moldea Nuestro Destino Biológico y las Implicaciones para la Salud, el Comportamiento y el Futuro de la InvestigaciónDe EverandMás allá del ADN: La Revolución Epigenética: Desde Mecanismos Celulares hasta Factores Ambientales: Cómo la Epigenética Moldea Nuestro Destino Biológico y las Implicaciones para la Salud, el Comportamiento y el Futuro de la InvestigaciónAún no hay calificaciones
- Genetica EvolutivaDocumento17 páginasGenetica EvolutivaIndira RamírezAún no hay calificaciones
- Genetica Mendliana 2013Documento67 páginasGenetica Mendliana 2013Indira RamírezAún no hay calificaciones
- Kevin PDFDocumento1 páginaKevin PDFIndira RamírezAún no hay calificaciones
- Kevin PDFDocumento1 páginaKevin PDFIndira RamírezAún no hay calificaciones
- Transferencia Embrionaria PorcinosDocumento9 páginasTransferencia Embrionaria PorcinosJohanita ArangoAún no hay calificaciones
- Cervicalgia Monografia (2) FEFEFEDocumento63 páginasCervicalgia Monografia (2) FEFEFEMaria Alvarez ParadaAún no hay calificaciones
- Hiperplasia Epitelial FocalDocumento7 páginasHiperplasia Epitelial FocalDarwinMurguiaDAún no hay calificaciones
- Acidos NucléicosDocumento23 páginasAcidos NucléicosAlejandra100% (1)
- Guia Conectores PreposicionesDocumento5 páginasGuia Conectores PreposicionesDagoberto Benítez MarínAún no hay calificaciones
- Tabla de NeoplasiasDocumento10 páginasTabla de NeoplasiasZhadow TorchonAún no hay calificaciones
- Asma BronquialDocumento23 páginasAsma BronquialJesus Alexis VladimirAún no hay calificaciones
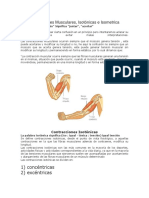
- Las Contracciones MuscularesDocumento3 páginasLas Contracciones MuscularessebastianAún no hay calificaciones
- Cuadro de MatrizDocumento2 páginasCuadro de MatrizBren AAún no hay calificaciones
- 1 US Marcadores Ecograficos de La Semana 18 A 22 OBSTETRICIADocumento15 páginas1 US Marcadores Ecograficos de La Semana 18 A 22 OBSTETRICIAEdery PulgarAún no hay calificaciones
- Histologia ReproductorDocumento5 páginasHistologia ReproductorMaría de los Angeles PerezAún no hay calificaciones
- Cuestionario de Las ExposicionesDocumento4 páginasCuestionario de Las ExposicionesMelanie LopezAún no hay calificaciones
- Silabo Morfo 2 PDFDocumento14 páginasSilabo Morfo 2 PDFdanielAún no hay calificaciones
- Proyecto de Investigacion (Ela)Documento26 páginasProyecto de Investigacion (Ela)mariennyanez100% (1)
- Bioquímica EspecialDocumento9 páginasBioquímica EspecialEdwardSerranoAún no hay calificaciones
- ProtozoariosDocumento17 páginasProtozoariosBrenda Eli QuispeAún no hay calificaciones
- Toma de Muestras de AguaDocumento21 páginasToma de Muestras de AguaSantosSimonMichelAún no hay calificaciones
- Reporte de La Titulación de ArgininaDocumento7 páginasReporte de La Titulación de ArgininaTommy Chente MartinezAún no hay calificaciones
- Trabajo Completo de CriminalisticaDocumento19 páginasTrabajo Completo de Criminalisticaanon_184232305Aún no hay calificaciones
- Ponton Falso 6.00 MDocumento2 páginasPonton Falso 6.00 MNebt HetAún no hay calificaciones
- HipoxiaDocumento25 páginasHipoxiaTony Vásquez Bautista100% (2)
- Almidon y Harina de Quinua y Cañihua Tesis UnaDocumento147 páginasAlmidon y Harina de Quinua y Cañihua Tesis Unafrancia lisbethAún no hay calificaciones
- Anexo Pef2016Documento148 páginasAnexo Pef2016AsesComDdigAún no hay calificaciones
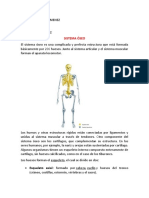
- Sistema Oseo, Muscular y La Piel AnatomíaDocumento19 páginasSistema Oseo, Muscular y La Piel AnatomíaIsabella Viveros JimenezAún no hay calificaciones
- Clinical Case 31-05-22Documento77 páginasClinical Case 31-05-22Ulises Hernández DávalosAún no hay calificaciones
- Sistema CardiovascularDocumento31 páginasSistema CardiovascularMarcos Yael PedrazaAún no hay calificaciones
- Diferencias Entre La Celula Animal y La Celula VegetalDocumento3 páginasDiferencias Entre La Celula Animal y La Celula VegetalPaul Andres Ruiz PantojaAún no hay calificaciones
- Teorías y Sistemas de La Psicología Semana2 PDocumento5 páginasTeorías y Sistemas de La Psicología Semana2 PENNERY ALEXANDREAún no hay calificaciones
- Factores Que Influyen en La CorrosionDocumento4 páginasFactores Que Influyen en La CorrosionAlberto Lopez0% (3)
- Que Es El BisturiDocumento6 páginasQue Es El BisturiMiguel Stick Peñaloza SanchezAún no hay calificaciones