También podría gustarte
- DESBROCE Y LIMPIEZA EN CARRETERAS Sección 201Documento7 páginasDESBROCE Y LIMPIEZA EN CARRETERAS Sección 201Italo Syb OR50% (2)
- Actividad para Trabajar CLASE MEMBRANA Y TRANSPORTEDocumento8 páginasActividad para Trabajar CLASE MEMBRANA Y TRANSPORTECamilo andres mejia vasquezAún no hay calificaciones
- Elena de White - El Pecado OcultoDocumento2 páginasElena de White - El Pecado OcultoSilvio MarianiAún no hay calificaciones
- Transferencia de MasaDocumento21 páginasTransferencia de Masa.:("*"BLacK BuLLeT"*"):.100% (3)
- Práctica 3 - PERMEABILIDAD CELULARDocumento11 páginasPráctica 3 - PERMEABILIDAD CELULARAnghelloLozanoDíaz100% (1)
- Osmosis y Permeabilidad Del Eritrocito (ResumenDocumento4 páginasOsmosis y Permeabilidad Del Eritrocito (Resumengiuseppegnr100% (4)
- Cuestionarios TodosDocumento48 páginasCuestionarios TodosSeeylin MejiaAún no hay calificaciones
- Guía Crossover PasivoDocumento4 páginasGuía Crossover Pasivofredyalz100% (1)
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017De EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Aún no hay calificaciones
- Transporte Pasivo, Osmosis, ExocitosisDocumento8 páginasTransporte Pasivo, Osmosis, ExocitosisAndres Higuera AceroAún no hay calificaciones
- Osmosis en Celulas de ElodeaDocumento2 páginasOsmosis en Celulas de Elodealaura moreno100% (1)
- GUIA DE PRACTICA de ÓSMOSIS Y PLASMÓLISIS EN PLANTAS FRAGILIDAD OSOMÓTICA ERITROCITOSDocumento7 páginasGUIA DE PRACTICA de ÓSMOSIS Y PLASMÓLISIS EN PLANTAS FRAGILIDAD OSOMÓTICA ERITROCITOSJavier Augusto Rodríguez Bravo100% (1)
- Los 10 Apps de Matemáticas para El TabletDocumento3 páginasLos 10 Apps de Matemáticas para El TabletbladicitoAún no hay calificaciones
- Presion Osmotica1Documento4 páginasPresion Osmotica1Jessy Rivaz HerreraAún no hay calificaciones
- Lectura 8Documento6 páginasLectura 8DANIEL ALEJANDROAún no hay calificaciones
- Acu A PorinasDocumento5 páginasAcu A PorinasChristian Fernando Rodriguez JimenesAún no hay calificaciones
- OsmolaridadDocumento3 páginasOsmolaridadErick Eduardo HernandezAún no hay calificaciones
- Guia Seminarios y TP Bio 379 - 2c I Semestre 2013Documento51 páginasGuia Seminarios y TP Bio 379 - 2c I Semestre 2013Francisca Mantaraya67% (6)
- Transporte de Agua A Través de Membranas BiológicasDocumento6 páginasTransporte de Agua A Través de Membranas BiológicasMatias PazAún no hay calificaciones
- Bracho R.T.Documento12 páginasBracho R.T.Roberto PazAún no hay calificaciones
- Balance Hidromineral y Equilibrio Acido-BaseDocumento12 páginasBalance Hidromineral y Equilibrio Acido-BaseVEROAún no hay calificaciones
- Trabajo BiologíaDocumento22 páginasTrabajo BiologíaNaylenAún no hay calificaciones
- Informe 5Documento8 páginasInforme 5Ruby Estefany García OrtízAún no hay calificaciones
- Practica Clase 2 PDFDocumento6 páginasPractica Clase 2 PDFCarlos RamosAún no hay calificaciones
- ReporteMembrana FisiologiaDocumento16 páginasReporteMembrana FisiologiaJulieta Muñoz50% (2)
- Laboratorio de FisiologíaDocumento10 páginasLaboratorio de FisiologíaMariana HopeAún no hay calificaciones
- Práctica 4. - OsmosisDocumento4 páginasPráctica 4. - OsmosisÁngel RomeroAún no hay calificaciones
- 1 Transporte MembranaDocumento4 páginas1 Transporte MembranaCarlos GabrielAún no hay calificaciones
- Practica de Laboratorio 2Documento9 páginasPractica de Laboratorio 2andrea123456Aún no hay calificaciones
- Practica de Laboratorio 2Documento9 páginasPractica de Laboratorio 2andrea123456Aún no hay calificaciones
- Transporte de AguaDocumento3 páginasTransporte de AguaChristian MSAún no hay calificaciones
- Septima Sesión de Biologia MVZ 2022-IDocumento58 páginasSeptima Sesión de Biologia MVZ 2022-I05-VZ-HU-PAOLA ESTEFANI QUISPE INCHEAún no hay calificaciones
- Biología Práctica Informe 04 Grupo 03 Permeabilidad CelularDocumento11 páginasBiología Práctica Informe 04 Grupo 03 Permeabilidad CelularEdwin GonzalesAún no hay calificaciones
- UrreaDocumento10 páginasUrreaGary Steven BuitragoAún no hay calificaciones
- s11 Art - En.esDocumento9 páginass11 Art - En.esGustavo Herrera MontenegroAún no hay calificaciones
- Taller 8 OsmosisDocumento3 páginasTaller 8 OsmosisJuanCarlosOrozcoCaicedoAún no hay calificaciones
- Práctica 4. - OsmosisDocumento4 páginasPráctica 4. - OsmosisÁngel RomeroAún no hay calificaciones
- Proteinas Canales de SodioDocumento10 páginasProteinas Canales de SodioMelissaAún no hay calificaciones
- Práctica 1 Permeabilidad de La Membrana 2015-2Documento5 páginasPráctica 1 Permeabilidad de La Membrana 2015-2Manuel OrihuelaAún no hay calificaciones
- Taller Agua, Electrolitos y Equilibrio Acido BaseDocumento17 páginasTaller Agua, Electrolitos y Equilibrio Acido BaseLuis Felipe Caro CaroAún no hay calificaciones
- Trabalho 01Documento9 páginasTrabalho 01Esdras LimaAún no hay calificaciones
- Preinforme BioquimicaDocumento6 páginasPreinforme BioquimicaAlejandra Gutierrez SaavedraAún no hay calificaciones
- Practica 6 MembranasDocumento5 páginasPractica 6 MembranasCora SalazarAún no hay calificaciones
- Permeabilidad de MembranasDocumento12 páginasPermeabilidad de MembranasWendys Orozco Maestre75% (8)
- Osmosis y Difusion LaboratorioDocumento14 páginasOsmosis y Difusion LaboratorioSodvith Camargo0% (1)
- Practica OsmosisDocumento6 páginasPractica OsmosisJhonatan Santos MartínezAún no hay calificaciones
- Practica 6 Informe MembranasDocumento5 páginasPractica 6 Informe MembranasandresAún no hay calificaciones
- FISIOLOGÍA GENERAL TALLER No 1 RESUELTODocumento4 páginasFISIOLOGÍA GENERAL TALLER No 1 RESUELTOjosè Alejandro Sinitave BuitragoAún no hay calificaciones
- Las Membranas Biológicas y TransporteDocumento37 páginasLas Membranas Biológicas y Transportepaola diazAún no hay calificaciones
- OsmosisDocumento12 páginasOsmosisJuan RasgadoAún no hay calificaciones
- Transporte A Través de MembranasDocumento11 páginasTransporte A Través de MembranasjavierAún no hay calificaciones
- Práctica Transporte A Traves de La MembranDocumento9 páginasPráctica Transporte A Traves de La Membranmpbeltran1Aún no hay calificaciones
- Interacciones Entre La Celula y El Medio ExtracelularDocumento45 páginasInteracciones Entre La Celula y El Medio Extracelularmarilarios94Aún no hay calificaciones
- Ósmosis en La Menbrana Semipermeable Del HuevoDocumento3 páginasÓsmosis en La Menbrana Semipermeable Del HuevoLourdes SalinasAún no hay calificaciones
- Biologia Practicas 18Documento6 páginasBiologia Practicas 18yosmi0% (1)
- Osmosis en Globulos Rojos - DefinitivoDocumento4 páginasOsmosis en Globulos Rojos - DefinitivoDairon Andrés Machado AgudeloAún no hay calificaciones
- Resumen de Fisiologia 2do ParcialDocumento57 páginasResumen de Fisiologia 2do ParcialJose Alain Candelaria RomeroAún no hay calificaciones
- Práctica 10 TRANSPORTE EN LA MEMBRANA CELULARDocumento6 páginasPráctica 10 TRANSPORTE EN LA MEMBRANA CELULARLUIS JOEL LOAYZA LLERENAAún no hay calificaciones
- Membrana PlasmáticaDocumento29 páginasMembrana PlasmáticaJulian CepedaAún no hay calificaciones
- Guia #04Documento3 páginasGuia #04aidaAún no hay calificaciones
- Ósmosis InversaDocumento4 páginasÓsmosis InversaCarolinaAún no hay calificaciones
- Clase A Clase Semana 3.2Documento13 páginasClase A Clase Semana 3.2DragmxAún no hay calificaciones
- Osmosis TermiDocumento8 páginasOsmosis TermiKimberly Andrea Marmani AriasAún no hay calificaciones
- Fisiología - Líquidos CorporalesDocumento5 páginasFisiología - Líquidos CorporalesMariamt RiveroAún no hay calificaciones
- Trastornos Del Balance de Liquidos y ElectrolitosDocumento20 páginasTrastornos Del Balance de Liquidos y ElectrolitosCalucho FelipeAún no hay calificaciones
- Apunte de Salival Volumen Celular 2015Documento21 páginasApunte de Salival Volumen Celular 2015Juan Pablo VivasAún no hay calificaciones
- Selladores 2022Documento22 páginasSelladores 2022Juan Pablo VivasAún no hay calificaciones
- Guía de Ejercicios Señalización 2015Documento4 páginasGuía de Ejercicios Señalización 2015Juan Pablo VivasAún no hay calificaciones
- Koino Yokan: General (Sin Numerar) GA 451 $2800.00Documento1 páginaKoino Yokan: General (Sin Numerar) GA 451 $2800.00Juan Pablo VivasAún no hay calificaciones
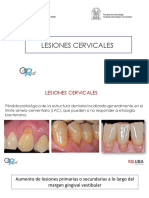
- Lesiones CervicalesDocumento64 páginasLesiones CervicalesJuan Pablo VivasAún no hay calificaciones
- Complejo Mayor de HistocompatibilidadDocumento2 páginasComplejo Mayor de HistocompatibilidadJuan Pablo VivasAún no hay calificaciones
- Venas Cara, Cuello y Torax PT.1Documento4 páginasVenas Cara, Cuello y Torax PT.1Juan Pablo VivasAún no hay calificaciones
- Ejercicios de Integración Clases 1 A 5Documento3 páginasEjercicios de Integración Clases 1 A 5Juan Pablo VivasAún no hay calificaciones
- Control Mecanico Del BiofilmDocumento17 páginasControl Mecanico Del BiofilmJuan Pablo VivasAún no hay calificaciones
- Registro de Estado DentarioDocumento23 páginasRegistro de Estado DentarioJuan Pablo VivasAún no hay calificaciones
- Antimicrobianos No Selectivos 2019Documento38 páginasAntimicrobianos No Selectivos 2019Juan Pablo VivasAún no hay calificaciones
- Cuestionario Lesiones ElementalesDocumento1 páginaCuestionario Lesiones ElementalesJuan Pablo VivasAún no hay calificaciones
- 1) 2018 CT1 AI Fisiologia de La Neurona y Neuroglia. IntroduccionDocumento36 páginas1) 2018 CT1 AI Fisiologia de La Neurona y Neuroglia. IntroduccionJuan Pablo VivasAún no hay calificaciones
- Anatomia Bioscopica Cavidad BucalDocumento4 páginasAnatomia Bioscopica Cavidad BucalJuan Pablo VivasAún no hay calificaciones
- 6G U4 MAT Sesion06Documento13 páginas6G U4 MAT Sesion06Castillo LuisAún no hay calificaciones
- Shertruck Pds EtchingprimerDocumento3 páginasShertruck Pds EtchingprimerAlvaro Mondragon BernalAún no hay calificaciones
- Practica 3 OriginalDocumento44 páginasPractica 3 Originalgladis bocanegra huamanAún no hay calificaciones
- Caso Clinico SX Wolff Parkinon WhiteDocumento20 páginasCaso Clinico SX Wolff Parkinon WhiteaylinAún no hay calificaciones
- 1 Lectura Evidencias para Identificar IonesDocumento3 páginas1 Lectura Evidencias para Identificar IonesmarisolAún no hay calificaciones
- 2015 Plan de Manejo Parque Nacional YbycuiDocumento193 páginas2015 Plan de Manejo Parque Nacional Ybycuirosa isabelAún no hay calificaciones
- Futuro UE-ICE PDFDocumento178 páginasFuturo UE-ICE PDFOrientadores PalenciaAún no hay calificaciones
- Pedagogía Social 5Documento9 páginasPedagogía Social 5comoestanamigos100% (2)
- NBX Manual Instruc Vimo 2-4-8Documento40 páginasNBX Manual Instruc Vimo 2-4-8Maru Ardusso100% (1)
- InfoTUB N 13-005 Cálculo de Espesor Del Aislamiento de Tuberías Plásticas Dic'13Documento6 páginasInfoTUB N 13-005 Cálculo de Espesor Del Aislamiento de Tuberías Plásticas Dic'13Nelly SuarezAún no hay calificaciones
- Homilia 20 de Marzo 2021Documento2 páginasHomilia 20 de Marzo 2021jaider hoyosAún no hay calificaciones
- Descargos Alberto Martinez - para Secretaria de EducacionDocumento56 páginasDescargos Alberto Martinez - para Secretaria de EducacionÁlvaro BarreraAún no hay calificaciones
- 08 - 0136 - EO Sistema de Telecontrol en Red de Media TensionDocumento132 páginas08 - 0136 - EO Sistema de Telecontrol en Red de Media Tensionsalmo83:18Aún no hay calificaciones
- Análisis de CasoDocumento3 páginasAnálisis de CasoL Carvallo Miguel0% (2)
- 2 Distribuciones de Frecuencia Unidad 2Documento17 páginas2 Distribuciones de Frecuencia Unidad 2Katherine TrianaAún no hay calificaciones
- LineaDocumento1 páginaLineatitorete100% (1)
- Trabajo N°5Documento4 páginasTrabajo N°5Cristian Lucas CuriAún no hay calificaciones
- Malla Curricular Especializacion Gerencia de Mercadeo PDFDocumento1 páginaMalla Curricular Especializacion Gerencia de Mercadeo PDFFabAún no hay calificaciones
- La Tierra en El UniversoDocumento10 páginasLa Tierra en El UniversoRosa MartinézAún no hay calificaciones
- VAIVAREJDocumento5 páginasVAIVAREJDaniel Scott McCorner VanKallisterAún no hay calificaciones
- TEO 7 2do P. Ortodoncia IIDocumento4 páginasTEO 7 2do P. Ortodoncia IIHtrebor EnihcebAún no hay calificaciones
- Conozcamos La BibliaDocumento21 páginasConozcamos La BibliajoseAún no hay calificaciones
- ReciboDigital 28022023Documento2 páginasReciboDigital 28022023Vladimir Santti FloresAún no hay calificaciones
- Ediciones Jurisprudencia Del Trabajo, C.A. RIF: J-00178041-6Documento32 páginasEdiciones Jurisprudencia Del Trabajo, C.A. RIF: J-00178041-6César CrespoAún no hay calificaciones
- Manual Drean Heladera DTP458BKRSS0Documento25 páginasManual Drean Heladera DTP458BKRSS0prodongoAún no hay calificaciones