También podría gustarte
- GuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisDe EverandGuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisCalificación: 4 de 5 estrellas4/5 (9)
- Informes 9 y 10-Grupo Jueves (Grupo 2) - Mesa 4Documento16 páginasInformes 9 y 10-Grupo Jueves (Grupo 2) - Mesa 4pabloAún no hay calificaciones
- Interpretación de análisis clínicos en perros y gatosDe EverandInterpretación de análisis clínicos en perros y gatosCalificación: 5 de 5 estrellas5/5 (2)
- Pruebas HematologicasDocumento7 páginasPruebas HematologicasLuigi OttoAún no hay calificaciones
- Hematología: Atlas de Morfología celularDe EverandHematología: Atlas de Morfología celularAún no hay calificaciones
- Cuadro Hematico CompletoDocumento19 páginasCuadro Hematico CompletoyancyAún no hay calificaciones
- Guía práctica para técnico superior de laboratorio de diagnóstico clínico y biomédicoDe EverandGuía práctica para técnico superior de laboratorio de diagnóstico clínico y biomédicoCalificación: 4.5 de 5 estrellas4.5/5 (10)
- Informe 2 de Analisis ClinicoDocumento20 páginasInforme 2 de Analisis ClinicoJuan Yarleque VilcaAún no hay calificaciones
- Rejuvenecer Con El Plasma Sanguíneo De Los JóvenesDe EverandRejuvenecer Con El Plasma Sanguíneo De Los JóvenesCalificación: 5 de 5 estrellas5/5 (1)
- Interpretacion Clinica Del Hemograma CompletoDocumento25 páginasInterpretacion Clinica Del Hemograma CompletoPercy VargasAún no hay calificaciones
- Marcadores de infecciones transmisibles vía transfusional: El caso del banco de sangre de la Escuela de Microbiología de la Universidad de Antioquia, 2015-2016De EverandMarcadores de infecciones transmisibles vía transfusional: El caso del banco de sangre de la Escuela de Microbiología de la Universidad de Antioquia, 2015-2016Calificación: 5 de 5 estrellas5/5 (1)
- Informe HemogramaDocumento9 páginasInforme HemogramaLiz Melissa VargasAún no hay calificaciones
- Clase 8. Anomalias HemogramaDocumento94 páginasClase 8. Anomalias HemogramaGUERSAN INGENIEROS S.R.LAún no hay calificaciones
- Introducción a la antropología biológica: Origen, variabilidad y adaptación de las poblaciones humanasDe EverandIntroducción a la antropología biológica: Origen, variabilidad y adaptación de las poblaciones humanasAún no hay calificaciones
- UABP 2. El Hemograma en La Práctica Clínica - Métodos Diagnósticos de Las Citopenias Inmunes - SangreDocumento21 páginasUABP 2. El Hemograma en La Práctica Clínica - Métodos Diagnósticos de Las Citopenias Inmunes - SangreMartín PanicoAún no hay calificaciones
- Hemograma en BovinosDocumento7 páginasHemograma en BovinosVictoria AguilarAún no hay calificaciones
- Coagulación Para Todos: Medicina Para TodosDe EverandCoagulación Para Todos: Medicina Para TodosCalificación: 4.5 de 5 estrellas4.5/5 (3)
- Hemograma OneDocumento9 páginasHemograma OneOneiscarGomezAún no hay calificaciones
- Métodos quelométricos y otros métodos volumétricos de análisis clínicosDe EverandMétodos quelométricos y otros métodos volumétricos de análisis clínicosAún no hay calificaciones
- Recuento de ReticulocitosDocumento8 páginasRecuento de ReticulocitosMarilyn CrissAún no hay calificaciones
- Informe HemogramaDocumento13 páginasInforme Hemogramamaria paula bejaranoAún no hay calificaciones
- La Biometría HemáticaDocumento4 páginasLa Biometría HemáticaBryan Edu0% (1)
- Hematología MicrobiologiaDocumento62 páginasHematología MicrobiologiaNegrita GuevaraAún no hay calificaciones
- ICSHDocumento102 páginasICSHMirco Galvez Hinostroza100% (1)
- Serie RojaDocumento31 páginasSerie RojaDavidCangasCenicerosAún no hay calificaciones
- Cimentaciones Safe Marcelo GuerraDocumento9 páginasCimentaciones Safe Marcelo GuerraDAVID CHATA PACOMPIAAún no hay calificaciones
- Hematograma PDFDocumento7 páginasHematograma PDFALEXA GABRIELA JIMENEZ RIOSAún no hay calificaciones
- Hematocrito BLANCODocumento6 páginasHematocrito BLANCOPedro Bejar QuispeAún no hay calificaciones
- Interpretacion de Examenes de LaboratorioDocumento23 páginasInterpretacion de Examenes de LaboratorioJocelyn MamaniAún no hay calificaciones
- Informe de Práctica 4Documento19 páginasInforme de Práctica 4Elias Charcape OtinianoAún no hay calificaciones
- Hemogram ADocumento62 páginasHemogram APaúl Andre MerloAún no hay calificaciones
- Serie BlancaDocumento14 páginasSerie BlancaTon SalmonAún no hay calificaciones
- Practica # 3 Recuento de Eritrocitos PDFDocumento4 páginasPractica # 3 Recuento de Eritrocitos PDFTamara VeraAún no hay calificaciones
- HEMATOLOGIADocumento5 páginasHEMATOLOGIAapi-3737581100% (3)
- Bioquimica Clinica-Sangre y OrinaDocumento23 páginasBioquimica Clinica-Sangre y OrinaGabriel BarzalloAún no hay calificaciones
- Evaluacion de Celulas Sanguine AsDocumento5 páginasEvaluacion de Celulas Sanguine AsRosa Maria Triana MejiaAún no hay calificaciones
- Parámetros HematimétricosDocumento15 páginasParámetros HematimétricosabbyCAún no hay calificaciones
- Interpretacion de Examenes de Laboratorio PDFDocumento21 páginasInterpretacion de Examenes de Laboratorio PDFJose100% (3)
- Interpretación HemogramaDocumento9 páginasInterpretación HemogramaSARA SANCHEZ BELTRANAún no hay calificaciones
- Interpretación Del Cuadro Hemático. Grupo 2. Microbiología. III Semestre UQ.Documento11 páginasInterpretación Del Cuadro Hemático. Grupo 2. Microbiología. III Semestre UQ.Jacobo GutierrezAún no hay calificaciones
- Celulas SanguineasDocumento11 páginasCelulas SanguineasCristhian Jaico CruzAún no hay calificaciones
- Informe 7-HematrimetriaDocumento35 páginasInforme 7-HematrimetriaDavid Villalobos BardalesAún no hay calificaciones
- Practica 2 Analisi (1) .Documento20 páginasPractica 2 Analisi (1) .Teresa Saavedra100% (2)
- PRACTICA ReticulocitosDocumento11 páginasPRACTICA ReticulocitosFATIMA ALINE IPATZI HERNANDEZAún no hay calificaciones
- Hematology Without The Numbers in Clinic Blood Film Evaluation - En.esDocumento22 páginasHematology Without The Numbers in Clinic Blood Film Evaluation - En.esMayra CifuentesAún no hay calificaciones
- Cuenta de Eritrocitos, Retis e Índices EritrocitariosDocumento12 páginasCuenta de Eritrocitos, Retis e Índices EritrocitariosKariana NóperiAún no hay calificaciones
- Laboratorio de UrgenciasDocumento4 páginasLaboratorio de UrgenciasJuan Carlos VázquezAún no hay calificaciones
- Hematocrito y Biometría HemáticaDocumento5 páginasHematocrito y Biometría HemáticaAlexandra medina100% (1)
- Presentación Hemato.Documento12 páginasPresentación Hemato.Maria Cruz BlassAún no hay calificaciones
- Hem U3 Cris Ortegon OsorioDocumento9 páginasHem U3 Cris Ortegon OsorioCris OrtegonAún no hay calificaciones
- Laboratorio Hemograma 2018Documento7 páginasLaboratorio Hemograma 2018Juan Carlos MéndezAún no hay calificaciones
- Unidad III Hemograma Su Interpretacion y Algoritmos de DX AnemiasDocumento46 páginasUnidad III Hemograma Su Interpretacion y Algoritmos de DX AnemiasariahopeeAún no hay calificaciones
- Anexo TP10Documento11 páginasAnexo TP10griselAún no hay calificaciones
- Practica 3 HemogramaDocumento63 páginasPractica 3 HemogramaJoseline PintoAún no hay calificaciones
- Informe 3-VALOR HEMATOCRITODocumento3 páginasInforme 3-VALOR HEMATOCRITOCindy Villacis0% (1)
- Indices EritrocitariosDocumento13 páginasIndices EritrocitariosScarleth Isabel Dávila HernándezAún no hay calificaciones
- Recuento de Eritrocitos PracticaDocumento5 páginasRecuento de Eritrocitos PracticakathyAún no hay calificaciones
- Recuento de ReticulocitosDocumento8 páginasRecuento de ReticulocitosQUIMICO CLINICO WILLIANS SANCHEZ100% (3)
- SanidadDocumento9 páginasSanidadEuge SanabriaAún no hay calificaciones
- Dimensionamiento de Una GranjaDocumento3 páginasDimensionamiento de Una GranjaEuge SanabriaAún no hay calificaciones
- Manejo de Cerdas en Bandas.Documento9 páginasManejo de Cerdas en Bandas.Euge SanabriaAún no hay calificaciones
- 1-Introducción - Estadísticas 2013Documento50 páginas1-Introducción - Estadísticas 2013Euge SanabriaAún no hay calificaciones
- Cronometría DentariaDocumento60 páginasCronometría DentariaEuge SanabriaAún no hay calificaciones
- Citologia Diagnòstica 2017Documento19 páginasCitologia Diagnòstica 2017Euge SanabriaAún no hay calificaciones
- Temario y Bibliografía-Secundaria Agraria-Directoras y DirectoresDocumento13 páginasTemario y Bibliografía-Secundaria Agraria-Directoras y DirectoresEuge SanabriaAún no hay calificaciones
- El Desarrollo Del Pollito BB Dia A DiaDocumento25 páginasEl Desarrollo Del Pollito BB Dia A DiaEuge SanabriaAún no hay calificaciones
- Carpeta ConejosDocumento65 páginasCarpeta ConejosEuge SanabriaAún no hay calificaciones
- El TamboDocumento13 páginasEl TamboEuge SanabriaAún no hay calificaciones
- Tema 15 - Alteraciones de Las PlaquetasDocumento4 páginasTema 15 - Alteraciones de Las PlaquetasClaudia Alonso PérezAún no hay calificaciones
- Examen 1º TRIMESTRE FISIOPATOLOGÍA GENERALDocumento12 páginasExamen 1º TRIMESTRE FISIOPATOLOGÍA GENERALSonia OlveraAún no hay calificaciones
- Cuestionario de BiologiaDocumento16 páginasCuestionario de BiologiaBetty PostigoAún no hay calificaciones
- Presentación Completa - Sem4Documento75 páginasPresentación Completa - Sem4Lucia InfinitivelifeAún no hay calificaciones
- Sistema MuscularDocumento27 páginasSistema MuscularTiare AnaisAún no hay calificaciones
- Tema 30 SasDocumento6 páginasTema 30 SasMaria RamirezAún no hay calificaciones
- Masajes para La Secuencia DorsalDocumento3 páginasMasajes para La Secuencia DorsalJesús Rodríguez MulatoAún no hay calificaciones
- Edicion Medlab Jul-SepDocumento19 páginasEdicion Medlab Jul-SepClaudia Naomi Ventura OrtizAún no hay calificaciones
- Psicología Pantera de Itzel Díaz - Psicoanálisis Del Rey LeónDocumento10 páginasPsicología Pantera de Itzel Díaz - Psicoanálisis Del Rey LeónYulimar TorresAún no hay calificaciones
- Aminoácidos. Equipo 2Documento2 páginasAminoácidos. Equipo 2FridaAún no hay calificaciones
- Fisiologia MuscularDocumento4 páginasFisiologia MuscularAngie Ochoa RealesAún no hay calificaciones
- Agenda - Bii - Biologìa IiDocumento3 páginasAgenda - Bii - Biologìa Iijosmargamer212Aún no hay calificaciones
- Pesos y Medidas de Los Órganos Del Cuerpo HumanoDocumento5 páginasPesos y Medidas de Los Órganos Del Cuerpo HumanoVeliaAún no hay calificaciones
- Mapa Sistema NerviosoDocumento7 páginasMapa Sistema NerviosoAlvarez Salas Frida XailAún no hay calificaciones
- Espacios PerifaringeosDocumento7 páginasEspacios PerifaringeosKelma GuimaraesAún no hay calificaciones
- Análisis Resultado Valores de Referencia Unidad: Somatomedina C (Igf-I)Documento1 páginaAnálisis Resultado Valores de Referencia Unidad: Somatomedina C (Igf-I)alejandro RuedaAún no hay calificaciones
- Sodio y PotasioDocumento2 páginasSodio y PotasioMilena FloresAún no hay calificaciones
- Diapositivas ArticulacionesDocumento39 páginasDiapositivas ArticulacionesEmpty AnaAún no hay calificaciones
- Apuntes, Hematologia LabDocumento3 páginasApuntes, Hematologia LabJuan David Cruz BerasAún no hay calificaciones
- Teoría-Función Cardiovascular - MEHU 632Documento24 páginasTeoría-Función Cardiovascular - MEHU 632Sara ParedesAún no hay calificaciones
- Enfoque CognitivoDocumento164 páginasEnfoque CognitivoCAMILA ELISABETH CHINGAL AYALAAún no hay calificaciones
- Embrio Tarea Academica 5Documento6 páginasEmbrio Tarea Academica 5Angelo CastilloAún no hay calificaciones
- Sistema Respiratorio Trabajo GrupalDocumento10 páginasSistema Respiratorio Trabajo GrupalGeorgina VillegasAún no hay calificaciones
- Fisiopatologia Cardiovascular CompendioDocumento47 páginasFisiopatologia Cardiovascular CompendioAlexandra AraujoAún no hay calificaciones
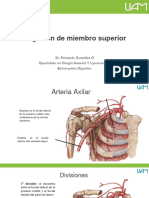
- Irrigacion Miembro SuperiorDocumento49 páginasIrrigacion Miembro Superiorcalebrayo16100% (1)
- Lipidos y MetabolismoDocumento45 páginasLipidos y Metabolismokona chanAún no hay calificaciones
- Cuestionario Tema 6 Metabolismo de Ácidos NucleicosDocumento21 páginasCuestionario Tema 6 Metabolismo de Ácidos NucleicosLiliana EscobarAún no hay calificaciones
- Anticuerpo F4 - 80Documento1 páginaAnticuerpo F4 - 80Leon NikolaiAún no hay calificaciones
- Funciones Sensoriales - Sistema SematosensorialDocumento8 páginasFunciones Sensoriales - Sistema SematosensorialAndrea Bulnes MatamorosAún no hay calificaciones
- Biología PDFDocumento2 páginasBiología PDFaikoowo kmcAún no hay calificaciones
- Altas Dosis: Los efectos milagrosos de dosis extremadamente altas de vitamina D3. El gran secreto que la industria farmacéutica quiere ocultarle.De EverandAltas Dosis: Los efectos milagrosos de dosis extremadamente altas de vitamina D3. El gran secreto que la industria farmacéutica quiere ocultarle.Calificación: 5 de 5 estrellas5/5 (4)
- Muchas Vidas, Muchos Sabios (Many Lives, Many Masters): (Many Lives, Many Masters)De EverandMuchas Vidas, Muchos Sabios (Many Lives, Many Masters): (Many Lives, Many Masters)Calificación: 4 de 5 estrellas4/5 (475)
- 50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaDe Everand50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaCalificación: 4 de 5 estrellas4/5 (37)
- Terapia cognitiva: Conceptos básicos y profundizaciónDe EverandTerapia cognitiva: Conceptos básicos y profundizaciónCalificación: 5 de 5 estrellas5/5 (1)
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- La metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceDe EverandLa metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceCalificación: 5 de 5 estrellas5/5 (8)
- GuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisDe EverandGuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisCalificación: 4 de 5 estrellas4/5 (9)
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- Sana tu Cuerpo, Calma tu Mente: Desintoxicar Hígado, Intestino Permeable, Salud Hormonal, Curación Emocional, Relajación, Ansiedad y Sanidad mental, Atención Plena, Psicoterapia y NutriciónDe EverandSana tu Cuerpo, Calma tu Mente: Desintoxicar Hígado, Intestino Permeable, Salud Hormonal, Curación Emocional, Relajación, Ansiedad y Sanidad mental, Atención Plena, Psicoterapia y NutriciónCalificación: 5 de 5 estrellas5/5 (4)
- Cardiología y enfermedades cardiovascularesDe EverandCardiología y enfermedades cardiovascularesCalificación: 5 de 5 estrellas5/5 (1)
- Notas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesDe EverandNotas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesAún no hay calificaciones
- Shiatsu: Conozca las técnicas y tratamientos de una de las terapias más antiguasDe EverandShiatsu: Conozca las técnicas y tratamientos de una de las terapias más antiguasAún no hay calificaciones
- Neuropsicología: Los fundamentos de la materiaDe EverandNeuropsicología: Los fundamentos de la materiaCalificación: 5 de 5 estrellas5/5 (1)
- Psiconeuroinmunología para la práctica clínicaDe EverandPsiconeuroinmunología para la práctica clínicaCalificación: 5 de 5 estrellas5/5 (4)
- El cerebro del niño explicado a los padresDe EverandEl cerebro del niño explicado a los padresCalificación: 4.5 de 5 estrellas4.5/5 (147)
- Abrázame fuerte: Siete conversaciones para un amor duraderoDe EverandAbrázame fuerte: Siete conversaciones para un amor duraderoManu BerásteguiCalificación: 4.5 de 5 estrellas4.5/5 (13)
- TDAH en Adultos. Cómo Reconocer y Tratar a un Adulto con TDAH en 30 Fáciles PasosDe EverandTDAH en Adultos. Cómo Reconocer y Tratar a un Adulto con TDAH en 30 Fáciles PasosCalificación: 4 de 5 estrellas4/5 (8)
- Fisiopatología de las enfermedades cardiovascularesDe EverandFisiopatología de las enfermedades cardiovascularesCalificación: 5 de 5 estrellas5/5 (1)
- Psicópatas seriales: Un recorrido por su oscura e inquietante naturalezaDe EverandPsicópatas seriales: Un recorrido por su oscura e inquietante naturalezaCalificación: 4 de 5 estrellas4/5 (3)
- Nuestra mente nos engaña: Sesgos y errores cognitivos que todos cometemosDe EverandNuestra mente nos engaña: Sesgos y errores cognitivos que todos cometemosCalificación: 3.5 de 5 estrellas3.5/5 (6)
- Dieta Para El Reflujo Biliar y Gastritis Alcalina - Incluye 20 Deliciosas Recetas Libres de Gluten y de Lácteos Para Tratar y Aliviar el Reflujo Biliar y Sus Molestos SíntomasDe EverandDieta Para El Reflujo Biliar y Gastritis Alcalina - Incluye 20 Deliciosas Recetas Libres de Gluten y de Lácteos Para Tratar y Aliviar el Reflujo Biliar y Sus Molestos SíntomasCalificación: 4 de 5 estrellas4/5 (9)
- Psicoterapia breve con niños y adolescentesDe EverandPsicoterapia breve con niños y adolescentesCalificación: 4.5 de 5 estrellas4.5/5 (15)
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)De EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Calificación: 5 de 5 estrellas5/5 (9)
- 200 tareas en terapia breve: 2ª ediciónDe Everand200 tareas en terapia breve: 2ª ediciónCalificación: 4.5 de 5 estrellas4.5/5 (33)
- ¿Cómo Prevenir O Curar El Cáncer?: ¡Conozca Los 3 Métodos Probados Contra El Cáncer!De Everand¿Cómo Prevenir O Curar El Cáncer?: ¡Conozca Los 3 Métodos Probados Contra El Cáncer!Aún no hay calificaciones