También podría gustarte
- Embriologia y Anatomia Ocular UNSAACDocumento36 páginasEmbriologia y Anatomia Ocular UNSAACJose Antonio Fuentes Vega100% (2)
- Distrofias VitreorretinianasDocumento10 páginasDistrofias VitreorretinianasrebelianoAún no hay calificaciones
- Taller 6 Biología Molecular - 201610 - MBIO1101 - 02 - .Documento14 páginasTaller 6 Biología Molecular - 201610 - MBIO1101 - 02 - .Jose David VelandiaAún no hay calificaciones
- Anatomía Del Nervio ÓpticoDocumento4 páginasAnatomía Del Nervio ÓpticoBladi RiveAún no hay calificaciones
- El CristalinoDocumento11 páginasEl CristalinoDr. jorge perez martinezAún no hay calificaciones
- Patologías de La CorneaDocumento10 páginasPatologías de La CorneaCarlos HernandezAún no hay calificaciones
- ACOMODACIÓNDocumento4 páginasACOMODACIÓNAgustin Ramirez100% (1)
- Embriologia Del Ojo 1 DownloableDocumento4 páginasEmbriologia Del Ojo 1 DownloableDayssi Maryll Valenzuela Manrique100% (1)
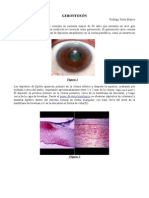
- Gerontoxón RABDocumento5 páginasGerontoxón RABRodrigo Avila BlancoAún no hay calificaciones
- Fotoquímica de La Visión Visión en Color ResumenDocumento3 páginasFotoquímica de La Visión Visión en Color Resumenlizzy rodriguez100% (1)
- Influencia de Las Ametropías en El Rendimiento Escolar en Niños de 9 A 12 Años en La Escuela Fiscal Esperanza Caputi OlveraDocumento13 páginasInfluencia de Las Ametropías en El Rendimiento Escolar en Niños de 9 A 12 Años en La Escuela Fiscal Esperanza Caputi Olverajuan100% (1)
- Glaucoma de Tensión NormalDocumento8 páginasGlaucoma de Tensión NormalCésar Augusto GarcíaAún no hay calificaciones
- VITREODocumento14 páginasVITREOREYES AGUILAR SALINAS100% (1)
- Uveitis AnteriorDocumento1 páginaUveitis AnteriorLuis Felipe MazariegosAún no hay calificaciones
- Clase Fisiologia CornealDocumento24 páginasClase Fisiologia CornealHugo FerreiraAún no hay calificaciones
- Fondo de OjoDocumento49 páginasFondo de OjoMaría Belén López Escalona100% (2)
- Anatomia Del Nervio OpticoDocumento6 páginasAnatomia Del Nervio OpticoAndres BenavidesAún no hay calificaciones
- Capas y Celulas de La RetinaDocumento2 páginasCapas y Celulas de La RetinaJonathan Álvarez Andrade100% (1)
- Guía Oftalmo 2° ParcialDocumento23 páginasGuía Oftalmo 2° Parcialmeremere1991Aún no hay calificaciones
- ParpadoDocumento43 páginasParpadokam estudio100% (1)
- Cuestionario OftalmosDocumento5 páginasCuestionario OftalmosMiguel CulmanAún no hay calificaciones
- Partes Lampara de HendiduraDocumento26 páginasPartes Lampara de HendiduraAgustin Ramirez0% (1)
- Vías VisualesDocumento4 páginasVías VisualesCesar Flores MorenoAún no hay calificaciones
- Anexos Del Globo Ocular y Medios RefringentesDocumento5 páginasAnexos Del Globo Ocular y Medios RefringentesAnibal Smith Santa Cruz Burga100% (1)
- Capas de La RetinaDocumento1 páginaCapas de La RetinaViviana MuñozAún no hay calificaciones
- Anexos OcularesDocumento4 páginasAnexos Ocularessarita escobarAún no hay calificaciones
- Bioquimica Iris y CCDocumento4 páginasBioquimica Iris y CCGianna LajaraAún no hay calificaciones
- Visión BinocularDocumento4 páginasVisión BinocularYui Dian Misa LizAún no hay calificaciones
- EsquematicoDocumento11 páginasEsquematicoDialexRobles100% (1)
- Av CartillasDocumento35 páginasAv CartillasFlores Santiago Axel Uriel100% (1)
- Taller Graf I CoDocumento5 páginasTaller Graf I CoNaywin Olascuaga TousAún no hay calificaciones
- Enfermedades de Los PárpadosDocumento12 páginasEnfermedades de Los PárpadosEnmanuel ValeroAún no hay calificaciones
- MIOPIADocumento23 páginasMIOPIAJose Antonio Ureta TrujilloAún no hay calificaciones
- Embriología de La CorneaDocumento7 páginasEmbriología de La Cornealuz mila sanchez100% (1)
- Pupila de Argyll RoberstonDocumento5 páginasPupila de Argyll RoberstonDrAgustin Hugo Alvarez AlvarezAún no hay calificaciones
- Tests Visión de ColorDocumento8 páginasTests Visión de Colorrodolfo gorveña100% (1)
- Clase Anisometropia y AniseiconiaDocumento41 páginasClase Anisometropia y Aniseiconiacrema236533100% (1)
- Fisiologia de La Visión 1Documento34 páginasFisiologia de La Visión 1Pris PerezAún no hay calificaciones
- Coloboma Del Nervio ÓpticoDocumento5 páginasColoboma Del Nervio ÓpticodovaAún no hay calificaciones
- Exploración Órbita y PárpadosDocumento42 páginasExploración Órbita y PárpadosJennifer MontoyaAún no hay calificaciones
- Expo QueratogloboDocumento22 páginasExpo QueratogloboKate CastañoAún no hay calificaciones
- ANISEICONÍADocumento2 páginasANISEICONÍACitlali Abigail Zamudio ZuñigaAún no hay calificaciones
- Anexos OcularesDocumento25 páginasAnexos OcularesMarvin O. Chuc UtrillaAún no hay calificaciones
- Ensayo - DEFECTOS REFRACTIVOS Y SU CORRECIONDocumento2 páginasEnsayo - DEFECTOS REFRACTIVOS Y SU CORRECIONDiego GomezAún no hay calificaciones
- Miopia y Sus ClasificacionesDocumento5 páginasMiopia y Sus ClasificacionesOctavioAún no hay calificaciones
- Enfermedades Del Cristalino PDFDocumento10 páginasEnfermedades Del Cristalino PDFFerdinand Ugarte MaqueraAún no hay calificaciones
- Guia de RetinosDocumento8 páginasGuia de RetinosZuleima DelgadoAún no hay calificaciones
- Exploración Vías PupilaresDocumento15 páginasExploración Vías PupilaresFelipe Alberto Naranjo HerreraAún no hay calificaciones
- Cristalino PDFDocumento98 páginasCristalino PDFAlma Rosa UGAún no hay calificaciones
- Semana 13. EstrabismoDocumento8 páginasSemana 13. EstrabismoIsa Cruz HipolitoAún no hay calificaciones
- CorneaDocumento6 páginasCorneaJoel BurgosAún no hay calificaciones
- ANTIGLAUCOMATOSOSDocumento2 páginasANTIGLAUCOMATOSOSmiguel luque lipaAún no hay calificaciones
- Aparato Lagrimal y Lagrima ITSCODocumento48 páginasAparato Lagrimal y Lagrima ITSCOJosePerez100% (1)
- Endotropia ExpoDocumento14 páginasEndotropia ExpoLuisferSamaniegoAún no hay calificaciones
- Aparato LagrimalDocumento13 páginasAparato LagrimalNicolas BaroneAún no hay calificaciones
- Copia de Historia Clinica OptometriaDocumento31 páginasCopia de Historia Clinica Optometriamaria alejandra garcia moncada100% (1)
- ExotropiasDocumento34 páginasExotropiasMarive24Aún no hay calificaciones
- CristalinoDocumento24 páginasCristalinoAura GarciaAún no hay calificaciones
- Ensayo Capitulo de Córnea - Mosby-Cristobal Rvera GreciaDocumento7 páginasEnsayo Capitulo de Córnea - Mosby-Cristobal Rvera GreciagreciaAún no hay calificaciones
- Histologia Del OjoDocumento16 páginasHistologia Del OjoAntony Carrero100% (1)
- Anatomía e Histología de La CórneaDocumento3 páginasAnatomía e Histología de La CórneaAndrea Mozo100% (1)
- Vacunas y PNI 2019Documento58 páginasVacunas y PNI 2019Luis F Concha Manriquez100% (1)
- Stents Medicados en Cardiología IntervencionistaDocumento8 páginasStents Medicados en Cardiología IntervencionistaLuis F Concha ManriquezAún no hay calificaciones
- Resumen MI Nefro Completo PDFDocumento25 páginasResumen MI Nefro Completo PDFLuis F Concha ManriquezAún no hay calificaciones
- Texto LlanosDocumento14 páginasTexto LlanosLuis F Concha ManriquezAún no hay calificaciones
- PP UssDocumento52 páginasPP UssLuis F Concha ManriquezAún no hay calificaciones
- Manual de Neo Puc 2012 JessyDocumento91 páginasManual de Neo Puc 2012 JessyLuis F Concha ManriquezAún no hay calificaciones
- Chile Crece ContigoDocumento17 páginasChile Crece ContigoLuis F Concha ManriquezAún no hay calificaciones
- FármacosDocumento4 páginasFármacosLuis F Concha Manriquez100% (1)
- Actividad Practica Embriologia Fecundacion-Segmentacion 2009Documento7 páginasActividad Practica Embriologia Fecundacion-Segmentacion 2009Nicolas Patricio SitjaAún no hay calificaciones
- Guía 1 4º BiologíaDocumento4 páginasGuía 1 4º BiologíaNintendo112Aún no hay calificaciones
- Practica Numero 3Documento8 páginasPractica Numero 3Anonymous phosYMdhRAún no hay calificaciones
- Crusigrama Biologia Segundo CorteDocumento2 páginasCrusigrama Biologia Segundo CorteSilvia Sandoval ContrerasAún no hay calificaciones
- Coagulograma 2020Documento6 páginasCoagulograma 2020Willy HuallpaAún no hay calificaciones
- Programación de Histología Embriología y GenéticaDocumento1 páginaProgramación de Histología Embriología y GenéticaSilva DennysAún no hay calificaciones
- Bioquímica Médica (4a. Ed.) - (18 Metabolismo de Las Lipoproteínas y Aterogénesis)Documento22 páginasBioquímica Médica (4a. Ed.) - (18 Metabolismo de Las Lipoproteínas y Aterogénesis)Ximena Miranda SalgadoAún no hay calificaciones
- 2 Estructura de LPPDocumento29 páginas2 Estructura de LPPRicardo CastilloAún no hay calificaciones
- ORGANELOS Exclusivos en CÉLULAS VEGETALESDocumento5 páginasORGANELOS Exclusivos en CÉLULAS VEGETALEScaled201185% (26)
- Lisosoma Endosoma PeroxisomaDocumento58 páginasLisosoma Endosoma Peroxisomatrilsol100% (1)
- Patologia RobbinsDocumento27 páginasPatologia RobbinsMaria Jose Cortes RodriguezAún no hay calificaciones
- Cascada de La CoagulaciónDocumento3 páginasCascada de La CoagulaciónEmely Rodríguez CastroAún no hay calificaciones
- Transporte A Través de La MembranaDocumento2 páginasTransporte A Través de La Membranacarlos BravoAún no hay calificaciones
- Actividad Tabla de Organelos CelularesDocumento2 páginasActividad Tabla de Organelos CelularesMasteremmm 10Aún no hay calificaciones
- Tipos de CicatrizaciónDocumento8 páginasTipos de CicatrizaciónTANIA CHAVEZ SANTILLANAún no hay calificaciones
- Celula Animal y VegealDocumento6 páginasCelula Animal y VegealMORENO MRCAún no hay calificaciones
- LIsosomas y PeroxisomasDocumento26 páginasLIsosomas y PeroxisomasVeronicajossimarAún no hay calificaciones
- Gametogénesis, Fecundación y Desarrollo Del Huevo Hasta La Formación de Las Tres Hojas Germinativas, ImplantaciónDocumento35 páginasGametogénesis, Fecundación y Desarrollo Del Huevo Hasta La Formación de Las Tres Hojas Germinativas, ImplantaciónKimberly CambisacaAún no hay calificaciones
- Muerte Celular UVMDocumento25 páginasMuerte Celular UVMAiramy AguilarAún no hay calificaciones
- Protocolo Microbiologia II MercedesDocumento32 páginasProtocolo Microbiologia II MercedesthchoperAún no hay calificaciones
- MITOSISDocumento6 páginasMITOSISCarlos Chambilla CastroAún no hay calificaciones
- Respuesta Inmune A ParasitosDocumento21 páginasRespuesta Inmune A ParasitosJuan MateusAún no hay calificaciones
- Unidad II Anatomia Y Fisiología de La CelulaDocumento26 páginasUnidad II Anatomia Y Fisiología de La Celulaana yuivi yucraAún no hay calificaciones
- BLAST ReporteDocumento13 páginasBLAST ReporteLily LeroAún no hay calificaciones
- Semestral San Marcos - Biología Semana 03Documento27 páginasSemestral San Marcos - Biología Semana 03Eduardo Huaroc FigueroaAún no hay calificaciones
- LÍPIDOS (Imprimir y Resolver Hoja 2 en El Cuaderno)Documento19 páginasLÍPIDOS (Imprimir y Resolver Hoja 2 en El Cuaderno)Maryam Deleon AlvaAún no hay calificaciones
- Dinámica MitocondrialDocumento19 páginasDinámica MitocondrialSandra Gissele Manzano GarcíaAún no hay calificaciones
- Taller Cinetica Enzimatica Sem A 2019Documento2 páginasTaller Cinetica Enzimatica Sem A 2019Alejandro MoralesAún no hay calificaciones
- Curacion de Heridas CutaneasDocumento16 páginasCuracion de Heridas CutaneasLizette RamirezAún no hay calificaciones