También podría gustarte
- Embarazo, parto y lactancia del bebéDe EverandEmbarazo, parto y lactancia del bebéAún no hay calificaciones
- Primera Semana Del FetoDocumento14 páginasPrimera Semana Del FetoJeff LAún no hay calificaciones
- 3 Ciclo UterinDocumento9 páginas3 Ciclo UterinAnonymous hkjp4qMVRsAún no hay calificaciones
- Fisiologia Gineco MarianDocumento14 páginasFisiologia Gineco Marianjanny Romero100% (1)
- Ciclo Mestrual, OvaricoDocumento32 páginasCiclo Mestrual, OvaricoCaro GuevaraAún no hay calificaciones
- Ciclo menstrual y ovárico enDocumento5 páginasCiclo menstrual y ovárico enAlessandro MoralesAún no hay calificaciones
- Trompas de Falopio y OvarioDocumento44 páginasTrompas de Falopio y OvarioAna LopezAún no hay calificaciones
- Norma Tecnica MujerDocumento9 páginasNorma Tecnica Mujerkarina cabanillasAún no hay calificaciones
- Ciclo OvaricoDocumento8 páginasCiclo OvaricoIka TapiaAún no hay calificaciones
- Citología Ginecológica Tema 2 RDocumento4 páginasCitología Ginecológica Tema 2 RJulieAún no hay calificaciones
- Aparato Reproductor FemeninoDocumento20 páginasAparato Reproductor FemeninoKoa NunchiAún no hay calificaciones
- PDF 20230816 171249 0000Documento44 páginasPDF 20230816 171249 0000Milena Jamileth Madrid VasquezAún no hay calificaciones
- Ciclo Ovárico y EndometrialDocumento5 páginasCiclo Ovárico y EndometrialAlma Saralena Cortez SanchesAún no hay calificaciones
- Gineco Parcial 1 Rec OrganizedDocumento232 páginasGineco Parcial 1 Rec OrganizedNaomy Us MartínezAún no hay calificaciones
- Histología Capitulo 22Documento3 páginasHistología Capitulo 22MarceloAisaAún no hay calificaciones
- Anatomía y Fisiología Del Aparato Reproductor de La YeguaDocumento11 páginasAnatomía y Fisiología Del Aparato Reproductor de La Yeguaelizabeht espa;aAún no hay calificaciones
- Manual Metodos Anticonceptivos (Servín Carrillo Axel Fernando)Documento30 páginasManual Metodos Anticonceptivos (Servín Carrillo Axel Fernando)Chavez Olivares Jade SarahiAún no hay calificaciones
- Aparato Reproductor FemeninoDocumento3 páginasAparato Reproductor FemeninoPaula TeijeiroAún no hay calificaciones
- Ciclo MenstrualDocumento47 páginasCiclo Menstrualbrian urreloAún no hay calificaciones
- SexualidadDocumento98 páginasSexualidadNataly Neira GuzmánAún no hay calificaciones
- Resumen Ross-Pawlina - Histología - Capitulo 23 - Sistema Genital FemeninoDocumento17 páginasResumen Ross-Pawlina - Histología - Capitulo 23 - Sistema Genital Femeninovictoriaetchegoyen123Aún no hay calificaciones
- Ciclo MenstrualDocumento22 páginasCiclo MenstrualAustinAún no hay calificaciones
- TAREA 10 Ciclo OvaricoDocumento40 páginasTAREA 10 Ciclo OvaricoCinthya Citlaly Sanchez CastroAún no hay calificaciones
- Ciclo SexualDocumento3 páginasCiclo Sexualsponjita28Aún no hay calificaciones
- Anatomía y fisiología del aparato reproductor femeninoDocumento25 páginasAnatomía y fisiología del aparato reproductor femeninoItan HCAún no hay calificaciones
- Embrio M1.4Documento15 páginasEmbrio M1.4Hector Efrain Garcia ChavezAún no hay calificaciones
- Ciclo MenstrualDocumento13 páginasCiclo MenstrualARIAS ROMERO EDUARDA NAOMYAún no hay calificaciones
- Ciclo Sexual FemeninoDocumento8 páginasCiclo Sexual FemeninoIngrid Daniela Campos LeañoAún no hay calificaciones
- CICLO OVÁRICO y EndometrialDocumento3 páginasCICLO OVÁRICO y EndometrialSelma Xiomara FragosoAún no hay calificaciones
- Gametogénesis, Ciclo Ovarico y Ciclo SexualDocumento7 páginasGametogénesis, Ciclo Ovarico y Ciclo SexualMaia ArraizAún no hay calificaciones
- 9 Resumen - Síntesis - Unidad 3 - Ross-Pawlina - Histología - Capitulo 23 - Sistema Genital Femenino (1)Documento17 páginas9 Resumen - Síntesis - Unidad 3 - Ross-Pawlina - Histología - Capitulo 23 - Sistema Genital Femenino (1)Miranda LeonettiAún no hay calificaciones
- Guia Embriologia CeabioDocumento5 páginasGuia Embriologia CeabioHaykareth Mi Amor BelloAún no hay calificaciones
- Ciclo Menstrual 7moDocumento23 páginasCiclo Menstrual 7moJENNIFER CORTESAún no hay calificaciones
- Ciclo MenstrualDocumento63 páginasCiclo MenstrualJimena Yucra JarroAún no hay calificaciones
- Trabajo 2 S. ReproductorDocumento9 páginasTrabajo 2 S. ReproductorMARTINA VÁSQUEZ BARRERAAún no hay calificaciones
- Útero y Ciclo MenstrualDocumento3 páginasÚtero y Ciclo MenstrualWismichu 4kAún no hay calificaciones
- Ciclo MestrualDocumento24 páginasCiclo MestrualLUIS LOZANOAún no hay calificaciones
- Aparato Reproductor FemeninoDocumento9 páginasAparato Reproductor FemeninoFuentes Perez ItzelAún no hay calificaciones
- Ciclo OvaricoDocumento13 páginasCiclo OvaricoEscarla FructuosoAún no hay calificaciones
- Fisiologia Del Ciclo MenstrualDocumento4 páginasFisiologia Del Ciclo MenstrualCristina RuizAún no hay calificaciones
- Desarrollo folicular y ovulación: del ciclo ovárico a la implantaciónDocumento12 páginasDesarrollo folicular y ovulación: del ciclo ovárico a la implantaciónSämael GabrielAún no hay calificaciones
- Aparato Reproductor MasculinoDocumento7 páginasAparato Reproductor MasculinoJosue ApazzaAún no hay calificaciones
- Fecundación, Historia.Documento11 páginasFecundación, Historia.Ivan R-Rosario67% (3)
- Anatomía femeninaDocumento82 páginasAnatomía femeninaHerimar Gonzalez PolancoAún no hay calificaciones
- Anat y Fisio Del Sist Rep Masc y Fem Parte 2Documento28 páginasAnat y Fisio Del Sist Rep Masc y Fem Parte 2Armando HernandezAún no hay calificaciones
- El Aparato Reproductor FemeninoDocumento5 páginasEl Aparato Reproductor FemeninoprototipoxzAún no hay calificaciones
- Resumen Cap 80 y 81 Fisiología Humana Medicina UNC FiladdDocumento1 páginaResumen Cap 80 y 81 Fisiología Humana Medicina UNC FiladdCamila AstudilloAún no hay calificaciones
- Anatomía y fisiología del aparato reproductor masculino y femeninoDocumento36 páginasAnatomía y fisiología del aparato reproductor masculino y femeninoluisis CaceresAún no hay calificaciones
- Hormonas y Ciclo Sexual Femenino y OvogenesisDocumento3 páginasHormonas y Ciclo Sexual Femenino y OvogenesisOscar Valdemar Hernandez LopezAún no hay calificaciones
- Semana 14 Fisiologia de Los Organos Internos FemeninoDocumento22 páginasSemana 14 Fisiologia de Los Organos Internos FemeninoAnghely Gianella Alicia Gomez MergildoAún no hay calificaciones
- Fisiología de ARF (Resumen)Documento6 páginasFisiología de ARF (Resumen)Christian Saúl Cruz AlonsoAún no hay calificaciones
- Ciclo MenstrualDocumento13 páginasCiclo MenstrualCarlos PernaleteAún no hay calificaciones
- Ciclo Ovárico y EndometrialDocumento4 páginasCiclo Ovárico y EndometrialAnahi AguileraAún no hay calificaciones
- Sistema Genital Femenino IDocumento63 páginasSistema Genital Femenino IValery ArdonAún no hay calificaciones
- Reproductor Femenino IDocumento9 páginasReproductor Femenino ITania Sofía Schaefer CzeraniukAún no hay calificaciones
- Histología del sistema reproductor femeninoDocumento16 páginasHistología del sistema reproductor femeninoKaisy Angelica Ramirez PeñaloAún no hay calificaciones
- Sistema Reproductor FemeninoDocumento7 páginasSistema Reproductor FemeninoEdglaxys VelasquezAún no hay calificaciones
- Resumen Fundamentos de Estructura y FunciónDocumento43 páginasResumen Fundamentos de Estructura y FunciónGénesis JiménezAún no hay calificaciones
- Sist. ReproductorDocumento36 páginasSist. Reproductorcamilabaez228Aún no hay calificaciones
- Potopeo Disertología ofDocumento9 páginasPotopeo Disertología ofVictorAún no hay calificaciones
- Vivir Mejor La MenopausiaDocumento199 páginasVivir Mejor La MenopausiaPaola Yanine Lopez Hernandez100% (2)
- Placenta PreviaDocumento33 páginasPlacenta PreviaMonse RodriguezAún no hay calificaciones
- PDF Aborto EspontaneoDocumento18 páginasPDF Aborto EspontaneoNatalia Fernandez PereiraAún no hay calificaciones
- Tarjetas EsiDocumento17 páginasTarjetas EsicamiisdAún no hay calificaciones
- Planificacion FamiliarDocumento42 páginasPlanificacion FamiliarJohan GómezAún no hay calificaciones
- Tabla Solo MisoprostolDocumento1 páginaTabla Solo MisoprostolAndrea RamirezAún no hay calificaciones
- Examen de Biologia Primer GradoDocumento3 páginasExamen de Biologia Primer GradoJUAN MANUEL GARCIA ORTEGAAún no hay calificaciones
- 01menopausia PDFDocumento9 páginas01menopausia PDFModa Para Ti-TalcaAún no hay calificaciones
- Cesarea - UpToDateDocumento19 páginasCesarea - UpToDateJRAAAún no hay calificaciones
- Descripción de Los Diferentes Estados en Vacas VacíasDocumento7 páginasDescripción de Los Diferentes Estados en Vacas VacíasGodofredo MamaniAún no hay calificaciones
- Acta de Inconcurrencia de CarpetaDocumento3 páginasActa de Inconcurrencia de CarpetaVanhia AguilarAún no hay calificaciones
- Qué es una doulaDocumento4 páginasQué es una doulaMorghannaAún no hay calificaciones
- Shock Hemorrágico UPAODocumento61 páginasShock Hemorrágico UPAOAlessandra Zapata CoronadoAún no hay calificaciones
- Exp de Genitales VagDocumento51 páginasExp de Genitales VagLucia BlaswongAún no hay calificaciones
- PropranololDocumento2 páginasPropranololMariana LopezAún no hay calificaciones
- GRR - Vaginitis Infecciosa PDFDocumento13 páginasGRR - Vaginitis Infecciosa PDFAlberto FloresAún no hay calificaciones
- Manual Lactancia UsuariosDocumento45 páginasManual Lactancia UsuariosDani Astudillo AceitónAún no hay calificaciones
- EVOLUCIÓN DE LA OBSTETRICIA EN EL MUNDODocumento8 páginasEVOLUCIÓN DE LA OBSTETRICIA EN EL MUNDOSianeAún no hay calificaciones
- Afiche Tecnicas de AmamantamientoDocumento1 páginaAfiche Tecnicas de AmamantamientoEstefi CastilloAún no hay calificaciones
- LISTA DE CHEQUEO No. 4 ATENCION DEL PARTODocumento4 páginasLISTA DE CHEQUEO No. 4 ATENCION DEL PARTOHillary Saray Carranza BatistaAún no hay calificaciones
- ANTICONCEPTIVOSDocumento17 páginasANTICONCEPTIVOSAndrea ChavezAún no hay calificaciones
- Control PrenatalDocumento3 páginasControl PrenatalKatherine LopezAún no hay calificaciones
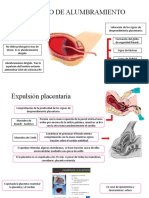
- Periodo de AlumbramientoDocumento3 páginasPeriodo de AlumbramientoPriscila100% (1)
- Ginecología y Su HistoriaDocumento22 páginasGinecología y Su HistoriandonaldluaccAún no hay calificaciones
- Parto VerticalDocumento49 páginasParto VerticalObstetra Ydelsa DelgadoAún no hay calificaciones
- Examen GinecológicoDocumento23 páginasExamen GinecológicoCristian Daniel Muñoz GómezAún no hay calificaciones
- Proyecto Del BebeDocumento9 páginasProyecto Del Bebeedson alayn muñoz quiñonesAún no hay calificaciones
- NST Leo FinalDocumento12 páginasNST Leo FinalLeonardoAún no hay calificaciones
- Detección temprana cáncer mamaDocumento34 páginasDetección temprana cáncer mamaRicardo Jesus Crespo MezaAún no hay calificaciones
- TDAH en Adultos. Cómo Reconocer y Tratar a un Adulto con TDAH en 30 Fáciles PasosDe EverandTDAH en Adultos. Cómo Reconocer y Tratar a un Adulto con TDAH en 30 Fáciles PasosCalificación: 4 de 5 estrellas4/5 (8)
- Terapia cognitiva: Conceptos básicos y profundizaciónDe EverandTerapia cognitiva: Conceptos básicos y profundizaciónCalificación: 5 de 5 estrellas5/5 (1)
- Trauma, miedo y amor: Hacia una autonomía interior con la ayuda de las constelacionesDe EverandTrauma, miedo y amor: Hacia una autonomía interior con la ayuda de las constelacionesCalificación: 1 de 5 estrellas1/5 (1)
- Psiconeuroinmunología para la práctica clínicaDe EverandPsiconeuroinmunología para la práctica clínicaCalificación: 5 de 5 estrellas5/5 (4)
- Fisiopatología de las enfermedades cardiovascularesDe EverandFisiopatología de las enfermedades cardiovascularesCalificación: 5 de 5 estrellas5/5 (1)
- La metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceDe EverandLa metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceCalificación: 5 de 5 estrellas5/5 (8)
- GuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisDe EverandGuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisCalificación: 4 de 5 estrellas4/5 (9)
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)De EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Calificación: 5 de 5 estrellas5/5 (9)
- El autismo: Reflexiones y pautas para comprenderlo y abordarloDe EverandEl autismo: Reflexiones y pautas para comprenderlo y abordarloCalificación: 4 de 5 estrellas4/5 (7)
- El libro conciso de la punción seca: Manual del terapeuta para las aplicaciones en los puntos gatillo miofasciales (Color)De EverandEl libro conciso de la punción seca: Manual del terapeuta para las aplicaciones en los puntos gatillo miofasciales (Color)Calificación: 3 de 5 estrellas3/5 (2)
- Ansiedad infantil. Los trastornos explicados a los padresDe EverandAnsiedad infantil. Los trastornos explicados a los padresCalificación: 4.5 de 5 estrellas4.5/5 (25)
- Prescripción de ejercico físico para la saludDe EverandPrescripción de ejercico físico para la saludCalificación: 5 de 5 estrellas5/5 (1)
- Magia Sexual - Tratado práctico de la ciencia oculta de los sexosDe EverandMagia Sexual - Tratado práctico de la ciencia oculta de los sexosCalificación: 4 de 5 estrellas4/5 (34)
- Psicoterapia breve con niños y adolescentesDe EverandPsicoterapia breve con niños y adolescentesCalificación: 4.5 de 5 estrellas4.5/5 (15)
- Plan De Dieta Para La Resistencia A La Insulina & Sistema Inmunológico En EspañolDe EverandPlan De Dieta Para La Resistencia A La Insulina & Sistema Inmunológico En EspañolCalificación: 3.5 de 5 estrellas3.5/5 (2)
- Póngase En Forma Sin Salir De Su CasaDe EverandPóngase En Forma Sin Salir De Su CasaCalificación: 4.5 de 5 estrellas4.5/5 (4)
- Puntos gatillo y cadenas musculares funcionales en osteopatía y terapia manual (Bicolor)De EverandPuntos gatillo y cadenas musculares funcionales en osteopatía y terapia manual (Bicolor)Calificación: 4.5 de 5 estrellas4.5/5 (23)
- Sana tu Cuerpo, Calma tu Mente: Desintoxicar Hígado, Intestino Permeable, Salud Hormonal, Curación Emocional, Relajación, Ansiedad y Sanidad mental, Atención Plena, Psicoterapia y NutriciónDe EverandSana tu Cuerpo, Calma tu Mente: Desintoxicar Hígado, Intestino Permeable, Salud Hormonal, Curación Emocional, Relajación, Ansiedad y Sanidad mental, Atención Plena, Psicoterapia y NutriciónCalificación: 5 de 5 estrellas5/5 (4)
- El código de la obesidad: Descifrando los secretos de la pérdida de pesoDe EverandEl código de la obesidad: Descifrando los secretos de la pérdida de pesoCalificación: 4.5 de 5 estrellas4.5/5 (51)
- Anatomía del caballo: Guía práctica ilustradaDe EverandAnatomía del caballo: Guía práctica ilustradaCalificación: 4 de 5 estrellas4/5 (9)
- La revolución de la tiroides y las glándulas suprarrenales: Un programa detallado para equilibrar tu metabolismo, cuidar tus hormonas y mejorar tu estado de ánimoDe EverandLa revolución de la tiroides y las glándulas suprarrenales: Un programa detallado para equilibrar tu metabolismo, cuidar tus hormonas y mejorar tu estado de ánimoCalificación: 4.5 de 5 estrellas4.5/5 (11)
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- Dieta Para El Reflujo Biliar y Gastritis Alcalina - Incluye 20 Deliciosas Recetas Libres de Gluten y de Lácteos Para Tratar y Aliviar el Reflujo Biliar y Sus Molestos SíntomasDe EverandDieta Para El Reflujo Biliar y Gastritis Alcalina - Incluye 20 Deliciosas Recetas Libres de Gluten y de Lácteos Para Tratar y Aliviar el Reflujo Biliar y Sus Molestos SíntomasCalificación: 4 de 5 estrellas4/5 (9)
- Kinesiotaping: Pruebas musculares y aplicaciones de taping (Color)De EverandKinesiotaping: Pruebas musculares y aplicaciones de taping (Color)Calificación: 5 de 5 estrellas5/5 (4)