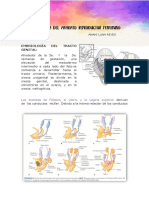
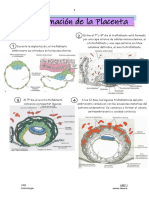
También podría gustarte
- Ginecología - 04 - Embriología Del Aparato Genital Femenino (Modificado)Documento57 páginasGinecología - 04 - Embriología Del Aparato Genital Femenino (Modificado)Hessler Hannsen Zambrano Condori100% (1)
- Embriología Del Aparato ReproductorDocumento83 páginasEmbriología Del Aparato ReproductorEdgardo Leal VillarroelAún no hay calificaciones
- AbortoDocumento13 páginasAbortoNikoll100% (1)
- Fisiología de La Menstruación PresentacionDocumento16 páginasFisiología de La Menstruación PresentacionDianny Garcia GarciaAún no hay calificaciones
- Fecundacinsegmentacineimplantacin 121206152556 Phpapp02Documento55 páginasFecundacinsegmentacineimplantacin 121206152556 Phpapp02María Esperanza Carrizo GarcíaAún no hay calificaciones
- Ciclo MenstrualDocumento22 páginasCiclo MenstrualPabloGonzalezAún no hay calificaciones
- Identidad Sexual y Roles de GeneroDocumento9 páginasIdentidad Sexual y Roles de GeneroAmanda ReyesAún no hay calificaciones
- Orientadoras 11 y 12 Morfofisiologia Humana 3Documento25 páginasOrientadoras 11 y 12 Morfofisiologia Humana 3Dayerlin HernandezAún no hay calificaciones
- Articulo Desarrollo Sexual Del EmbrionDocumento14 páginasArticulo Desarrollo Sexual Del EmbrionJeisson VecinoAún no hay calificaciones
- Desarrollo CompletoDocumento12 páginasDesarrollo CompletoVeronica AlexandraAún no hay calificaciones
- Anomalias Del Desarrollo SexualDocumento33 páginasAnomalias Del Desarrollo SexualjosheliinAún no hay calificaciones
- Material Complementario Desarrollo Embrionario Sistema Reproductor Femenino y MasculinoDocumento7 páginasMaterial Complementario Desarrollo Embrionario Sistema Reproductor Femenino y MasculinoEsmeraldaAún no hay calificaciones
- Diferenciación SexualDocumento11 páginasDiferenciación SexualDanni AlejaAún no hay calificaciones
- ConclusionesDocumento11 páginasConclusionesRosangelica Castro ParedesAún no hay calificaciones
- Anatomía e Histología de La Diferenciación Sexual EmbrioDocumento8 páginasAnatomía e Histología de La Diferenciación Sexual EmbrioVICENTE MOJICA RODRIGUEZAún no hay calificaciones
- Histo Semana 2Documento20 páginasHisto Semana 2HeydiAún no hay calificaciones
- Identidad SexualDocumento6 páginasIdentidad SexualjimenaAún no hay calificaciones
- Diferenciación Sexual Embrio-Fetal - de Las Moléculas A La AnatomiaDocumento11 páginasDiferenciación Sexual Embrio-Fetal - de Las Moléculas A La AnatomiaCinthia OchoaAún no hay calificaciones
- Capitulo 18 Embriogenesis Aparato Genital 2017Documento16 páginasCapitulo 18 Embriogenesis Aparato Genital 2017sandraAún no hay calificaciones
- Diferenciacion Sexual - EnsayoDocumento3 páginasDiferenciacion Sexual - EnsayoJessy BailonAún no hay calificaciones
- Diferenciación GonadalDocumento8 páginasDiferenciación GonadalLily Losada EstradaAún no hay calificaciones
- Dayana UrbinaDocumento2 páginasDayana UrbinaCarlos ConradoAún no hay calificaciones
- Genética Del Sexo y La ConductaDocumento11 páginasGenética Del Sexo y La ConductaSANAA BOUSOUARET BOUJEDDAINE-TSOULIAún no hay calificaciones
- Diferenciación Sexual y Embriología Del Aparato ReproductorDocumento19 páginasDiferenciación Sexual y Embriología Del Aparato ReproductorKoraima EstradaAún no hay calificaciones
- Síntesis PDFDocumento5 páginasSíntesis PDFJesica Flores SanluisAún no hay calificaciones
- Diferenciación Del SexoDocumento4 páginasDiferenciación Del SexoAndrea DelgadoAún no hay calificaciones
- Embriologia Del Aparato Reproductor Femenino y de La Mama 1204514323113786 3Documento28 páginasEmbriologia Del Aparato Reproductor Femenino y de La Mama 1204514323113786 3Gallardo RichardAún no hay calificaciones
- Embriología Del Sistema Genital FemeninoDocumento8 páginasEmbriología Del Sistema Genital FemeninoAldo IniestaAún no hay calificaciones
- Embriologia Hoy XDDocumento129 páginasEmbriologia Hoy XDfreshaAún no hay calificaciones
- Informe GametogenesisDocumento16 páginasInforme GametogenesisJonathan MejiaAún no hay calificaciones
- Wa0078.Documento9 páginasWa0078.Serrat MAún no hay calificaciones
- Sexo GeneticoDocumento5 páginasSexo GeneticoByron GuachisacaAún no hay calificaciones
- UNIDAD IX. Aparato GenitalDocumento23 páginasUNIDAD IX. Aparato GenitalcarlosmedanAún no hay calificaciones
- Diferenciacion Sexual y Patologica IiDocumento25 páginasDiferenciacion Sexual y Patologica IiFátima Coaguila MamaniAún no hay calificaciones
- Monografía ARFDocumento73 páginasMonografía ARFItzel Mosqueda RomeroAún no hay calificaciones
- 1Documento5 páginas1Mikaela lAún no hay calificaciones
- Desarrollo de La Sexualidad: Mg. Sandra P. GuillénDocumento31 páginasDesarrollo de La Sexualidad: Mg. Sandra P. GuillénLuis Gabriel Rubiano MoraAún no hay calificaciones
- Embriologia Del Aparato Reproductor Femenino-Anahi Luna ReyesDocumento7 páginasEmbriologia Del Aparato Reproductor Femenino-Anahi Luna ReyesAnahí LunaAún no hay calificaciones
- El Ovario Como Organo EndocrinoDocumento15 páginasEl Ovario Como Organo EndocrinoLAURA VALENTINA ALARCON RIOSAún no hay calificaciones
- MFH Iii - Ao 11Documento16 páginasMFH Iii - Ao 11Dianne RodilAún no hay calificaciones
- MFH Iii - Ao 11Documento16 páginasMFH Iii - Ao 11Escarleth CamejoAún no hay calificaciones
- Embriologia Del Aparato Reproductor Femenino y de La Mama 1204514323113786 3Documento28 páginasEmbriologia Del Aparato Reproductor Femenino y de La Mama 1204514323113786 3Drakko FernándezAún no hay calificaciones
- Manual Completo GinecologiaDocumento116 páginasManual Completo GinecologiaPdro SalvadorAún no hay calificaciones
- UNIDAD IV. Aparato GenitalDocumento32 páginasUNIDAD IV. Aparato GenitaljaviluzmAún no hay calificaciones
- Embriología Del Aparato Reproductor FemeninoDocumento4 páginasEmbriología Del Aparato Reproductor FemeninoCinthya VallesAún no hay calificaciones
- Embriología de GenitalesDocumento7 páginasEmbriología de GenitalesViviana PalaciosAún no hay calificaciones
- Informe Sobre Las Etapas Del Desarrollo EmbrionarioDocumento8 páginasInforme Sobre Las Etapas Del Desarrollo Embrionariopayalag232Aún no hay calificaciones
- Informe Sindrome Intersexual FemeninoDocumento8 páginasInforme Sindrome Intersexual FemeninoJordy AriasAún no hay calificaciones
- Embriologia y Anatomia Del Aparato Reproductor FemeninoDocumento3 páginasEmbriologia y Anatomia Del Aparato Reproductor FemeninoMiguel Correa CotuáAún no hay calificaciones
- Anatomía y Fisiología Ginecológica PanamericanaDocumento15 páginasAnatomía y Fisiología Ginecológica PanamericanaJaneth LeeAún no hay calificaciones
- Practica 13Documento3 páginasPractica 13Benjamín AlejandroAún no hay calificaciones
- Embrio FemDocumento3 páginasEmbrio FemValenAún no hay calificaciones
- Monografia - Sistema Genital - Correlaciones Clinicas-GDocumento10 páginasMonografia - Sistema Genital - Correlaciones Clinicas-GGabriella MonasterioAún no hay calificaciones
- GónadasDocumento14 páginasGónadasAguilar Callado Johari TadeaAún no hay calificaciones
- Desarrollo de Las Gonadas Masculinas y FemeninasDocumento3 páginasDesarrollo de Las Gonadas Masculinas y FemeninasAle AntonioAún no hay calificaciones
- Embriologia Del Aparato Genital FemeninoDocumento24 páginasEmbriologia Del Aparato Genital FemeninoAgustina milena Gonzalez AAún no hay calificaciones
- Informe Embriologia GenitalesDocumento6 páginasInforme Embriologia GenitalesAlejandro DuranAún no hay calificaciones
- Embriol. - Ap. Femenino - 18Documento19 páginasEmbriol. - Ap. Femenino - 18Escarleth Quevedo ChavezAún no hay calificaciones
- EmbriologiaDocumento10 páginasEmbriologiaboroboritoAún no hay calificaciones
- Genética y SexualidadDocumento9 páginasGenética y SexualidadHebert Jhimsy Pastor GarcíaAún no hay calificaciones
- Lecturas 5,6,7Documento32 páginasLecturas 5,6,7PEDRO MANUEL CASTRO RIVERAAún no hay calificaciones
- GametogenesisDocumento12 páginasGametogenesisDidi CHILLI'N BURRITO'SAún no hay calificaciones
- Informe de Invernadero de RosasDocumento14 páginasInforme de Invernadero de RosasFranz omar Jamanca GonzalesAún no hay calificaciones
- Fenologia en PaltoDocumento6 páginasFenologia en PaltoJuan Ccerhuayo HuamaniAún no hay calificaciones
- Mapas Conceptuales Ciclo Biológico CompletoDocumento9 páginasMapas Conceptuales Ciclo Biológico CompletoOMARAún no hay calificaciones
- Neurospora Crassa (Resumen)Documento2 páginasNeurospora Crassa (Resumen)Daysi LuAún no hay calificaciones
- Preguntas de RciuDocumento28 páginasPreguntas de Rciuchavitodata100% (2)
- Unidad #18Documento18 páginasUnidad #18Gilda MendezAún no hay calificaciones
- Ambiental 11-3Documento6 páginasAmbiental 11-3laura arizaAún no hay calificaciones
- Prep Citologia Vaginal FrotisDocumento2 páginasPrep Citologia Vaginal FrotisCamilo SiabatoAún no hay calificaciones
- Rep Sex Animales Formato - PPTX 1Documento1 páginaRep Sex Animales Formato - PPTX 1Vania GOAún no hay calificaciones
- Segunda Parte Proyecto 3 AñoDocumento2 páginasSegunda Parte Proyecto 3 AñoSary RojasAún no hay calificaciones
- El Genoma Humano y Las Bases Cromosómicas de La Herencia - RESUMENDocumento7 páginasEl Genoma Humano y Las Bases Cromosómicas de La Herencia - RESUMENValAún no hay calificaciones
- Ovogenesis Aporte SimpleDocumento5 páginasOvogenesis Aporte SimpleJavier Saldivia CardozoAún no hay calificaciones
- Progesterona 2da Generación Progestágenos SolosDocumento16 páginasProgesterona 2da Generación Progestágenos SolosKAREN VALERIA ROMERO ABADAún no hay calificaciones
- CICLO MENSTRUAL Angelo Duarte - 105359Documento22 páginasCICLO MENSTRUAL Angelo Duarte - 105359yessik pardoAún no hay calificaciones
- Catedra de GinecobstetriciaDocumento93 páginasCatedra de GinecobstetriciaHelen Aquino GarciaAún no hay calificaciones
- Tarea 2 - Seres VivosDocumento6 páginasTarea 2 - Seres VivosDavinson RodriguezAún no hay calificaciones
- Preguntas 6 7 8Documento11 páginasPreguntas 6 7 8Yordan Smith BustosAún no hay calificaciones
- Guia Virtual - Ciencias Naturales - Sexto GradoDocumento6 páginasGuia Virtual - Ciencias Naturales - Sexto GradoJohana MariaAún no hay calificaciones
- Placenta EmbrioUABP3Documento6 páginasPlacenta EmbrioUABP3Estrella LlenqueAún no hay calificaciones
- 8.-Reproduccion Sexual y AsexualDocumento5 páginas8.-Reproduccion Sexual y AsexualCarlos AlavaAún no hay calificaciones
- FitomejoramientoDocumento5 páginasFitomejoramientofernandohualAún no hay calificaciones
- Ciclo MenstrualDocumento20 páginasCiclo MenstrualKatherine Roncal MontalvoAún no hay calificaciones
- Plantas Madres, Selección de Explantes InicialesDocumento30 páginasPlantas Madres, Selección de Explantes InicialesLeandro ChilanAún no hay calificaciones
- Gonadas MasculinasDocumento10 páginasGonadas MasculinasEnory ReyesAún no hay calificaciones
- V o F SexualidadDocumento1 páginaV o F SexualidadmariaAún no hay calificaciones
- La FlorDocumento18 páginasLa FlorEdo tetragramatonAún no hay calificaciones