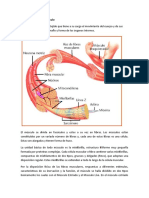
También podría gustarte
- Muscular y CardiovascularDocumento20 páginasMuscular y CardiovascularOctavio GarroneAún no hay calificaciones
- Lab FisioDocumento3 páginasLab FisioRosbelin PichardoAún no hay calificaciones
- PDF Las Interacciones de Las Celulas Entre Si Grupo 5 DDDocumento7 páginasPDF Las Interacciones de Las Celulas Entre Si Grupo 5 DDpilar morochoAún no hay calificaciones
- Tejido MuscularDocumento13 páginasTejido MuscularCamila Agostina VillafañeAún no hay calificaciones
- Biología Celular y Molecular Del MusculoDocumento11 páginasBiología Celular y Molecular Del MusculoMercy MoralesAún no hay calificaciones
- 9 - Citoesqueleto - Medicina 2021Documento14 páginas9 - Citoesqueleto - Medicina 2021Lomito CompletoAún no hay calificaciones
- Matriz Extracelular y Uniones Celulares 2011Documento28 páginasMatriz Extracelular y Uniones Celulares 2011Karla Labbe RodriguezAún no hay calificaciones
- Biología Celular Del MúsculoDocumento10 páginasBiología Celular Del MúsculoJuan MattosAún no hay calificaciones
- Histología Del Tejido Muscular y FisiologíaDocumento10 páginasHistología Del Tejido Muscular y FisiologíaDanna TanchivaAún no hay calificaciones
- Tema 5 AcabadoDocumento10 páginasTema 5 AcabadoMagdalena Oñate SerranoAún no hay calificaciones
- TEJIDOS RESUmen1Documento18 páginasTEJIDOS RESUmen1Aracely AdcAún no hay calificaciones
- Músculo: Repasando Un PocoDocumento16 páginasMúsculo: Repasando Un PocoLaru MarguliesAún no hay calificaciones
- 9 Conferencia Tejido MuscularDocumento101 páginas9 Conferencia Tejido MuscularHenry CondoriAún no hay calificaciones
- Tejido Muscular ÚltDocumento49 páginasTejido Muscular Últvideos MiosAún no hay calificaciones
- Cap 66Documento17 páginasCap 66André MaquinoAún no hay calificaciones
- Rae BQDocumento10 páginasRae BQMaria Victoria Martinez HernandezAún no hay calificaciones
- Clase - 7 Adhesión Celular-4 Dif.Documento28 páginasClase - 7 Adhesión Celular-4 Dif.KAREN VIVIANA PAVEZ RODRIGUEZAún no hay calificaciones
- Cito EsqueletoDocumento68 páginasCito EsqueletoDiego PantojaAún no hay calificaciones
- Wuolah Free Tema 6 y 7 Gulag FreeDocumento13 páginasWuolah Free Tema 6 y 7 Gulag FreealbaburcalAún no hay calificaciones
- Tema 2. Glicocalix y Especializaciones de MembranaDocumento4 páginasTema 2. Glicocalix y Especializaciones de MembranaMercedes MartínAún no hay calificaciones
- Guia Tejido MuscularDocumento6 páginasGuia Tejido MuscularDiana Esmeralda De los SantosAún no hay calificaciones
- Tema 6 BiocelDocumento8 páginasTema 6 Biocelrodriguezmartinezm022Aún no hay calificaciones
- CitoesqueletoDocumento6 páginasCitoesqueletoLoreto Santana SchwarzeneggerAún no hay calificaciones
- Soporte, Locomoción, Almacenamiento y Reciclado Celular PDFDocumento5 páginasSoporte, Locomoción, Almacenamiento y Reciclado Celular PDFJimena M. M.Aún no hay calificaciones
- Biologia Celular y Molecular de Robertis 16 Ed Primera ParteDocumento13 páginasBiologia Celular y Molecular de Robertis 16 Ed Primera ParteLilian NairAún no hay calificaciones
- Biologia 7Documento15 páginasBiologia 7Lou-LouAún no hay calificaciones
- Tejido MuscularDocumento34 páginasTejido MuscularRosa Marcela Rojas CorralesAún no hay calificaciones
- Citoesqueleto y Uniones CelularesDocumento22 páginasCitoesqueleto y Uniones CelularesGemma PrzAún no hay calificaciones
- Introducción Al CitoesqueletoDocumento5 páginasIntroducción Al CitoesqueletoAina cuixart gimoAún no hay calificaciones
- Capitulo 5 GanongDocumento9 páginasCapitulo 5 GanongMeli MolinaAún no hay calificaciones
- 7 Tejido - Muscular Ok 2020 - 1Documento11 páginas7 Tejido - Muscular Ok 2020 - 1Romi SAún no hay calificaciones
- Cap 6 6-6 y 6-7Documento4 páginasCap 6 6-6 y 6-7Adrimar CortésAún no hay calificaciones
- CitoesqueletoDocumento2 páginasCitoesqueletointeligenciaemocional003Aún no hay calificaciones
- Fisiologia de Del MusculoDocumento5 páginasFisiologia de Del Musculojeinnymar padronAún no hay calificaciones
- Músculo LisoDocumento13 páginasMúsculo Lisocepj1908Aún no hay calificaciones
- CitoesqueletoDocumento22 páginasCitoesqueletomaria aline rezende de souzaAún no hay calificaciones
- Tejido Muscula1Documento4 páginasTejido Muscula1Kristell Davila MartínezAún no hay calificaciones
- 3.1 Matriz Citoplasma - Tica y CitoesqueletoDocumento33 páginas3.1 Matriz Citoplasma - Tica y CitoesqueletoElvirita Vanessa PereiraAún no hay calificaciones
- Unidad 2-SISTEMA NEUROMUSCULAR SISTEMA NEUROMUSCULARDocumento20 páginasUnidad 2-SISTEMA NEUROMUSCULAR SISTEMA NEUROMUSCULARTomi CatalaAún no hay calificaciones
- Certamen III, Biología CelularDocumento27 páginasCertamen III, Biología CelularSinia DiazAún no hay calificaciones
- Tejido MuscularDocumento41 páginasTejido MuscularDeize SartoriAún no hay calificaciones
- Poblaciones CelularesDocumento20 páginasPoblaciones CelularesDionik CasasAún no hay calificaciones
- Tema 7 Uniones CelularesDocumento6 páginasTema 7 Uniones CelularesDavid Molina LópezAún no hay calificaciones
- Soporte, Locomoción, Almacenamiento y Reciclado CelularDocumento4 páginasSoporte, Locomoción, Almacenamiento y Reciclado CelularJimena M. M.100% (1)
- Estructura y Función MuscularDocumento2 páginasEstructura y Función Muscularcamilo jose rodriguez ayubiAún no hay calificaciones
- UNIDAD 5 CitoesqueletoDocumento58 páginasUNIDAD 5 CitoesqueletoCarolina Bravo BastidasAún no hay calificaciones
- Informe MuscularDocumento17 páginasInforme MuscularAlejandro AlvaradoAún no hay calificaciones
- Tejido MuscularDocumento4 páginasTejido MuscularSandra TorelliAún no hay calificaciones
- CitoesqueletoDocumento66 páginasCitoesqueletoBetty PostigoAún no hay calificaciones
- 12 Tejido MuscularDocumento8 páginas12 Tejido MuscularAldana MoralesAún no hay calificaciones
- Tejido Muscular TC ESPECIALIZADO 3°semana - 1Documento18 páginasTejido Muscular TC ESPECIALIZADO 3°semana - 1HERNAN GONZALEZAún no hay calificaciones
- F ActinaDocumento10 páginasF ActinaAgustina UriarteAún no hay calificaciones
- Semana 5 Clase 1 Tejido Muscular 2020Documento81 páginasSemana 5 Clase 1 Tejido Muscular 2020JHAMPIER QUISPE HUALLPAAún no hay calificaciones
- Tejido MuscularDocumento3 páginasTejido Muscular301Perez Pérez Daniela SofíaAún no hay calificaciones
- Apuntes Tercer ParcialDocumento78 páginasApuntes Tercer Parcialdservin20Aún no hay calificaciones
- Biologia Celular e Histología - Esther Fdez PDFDocumento114 páginasBiologia Celular e Histología - Esther Fdez PDFsaraAún no hay calificaciones
- Tema 3Documento7 páginasTema 3letiAún no hay calificaciones
- Tema 2 cONJUNTIVODocumento3 páginasTema 2 cONJUNTIVOvega45gaAún no hay calificaciones
- Aformularioregistro01 ImplDocumento2 páginasAformularioregistro01 ImplCarlos VargasAún no hay calificaciones
- Eventos Oficiales Folklore Mujeres Titulares y SuplentesDocumento6 páginasEventos Oficiales Folklore Mujeres Titulares y SuplentesCarlos VargasAún no hay calificaciones
- Simen Ascensores 17194Documento1 páginaSimen Ascensores 17194Carlos VargasAún no hay calificaciones
- Mant Gaston Faltantes-1Documento1 páginaMant Gaston Faltantes-1Carlos VargasAún no hay calificaciones
- Ups Ult 1000 3000VTDocumento2 páginasUps Ult 1000 3000VTCarlos VargasAún no hay calificaciones
- ManualUPSPolarisXION650 850Documento12 páginasManualUPSPolarisXION650 850Carlos Vargas100% (1)
- LABORATORIO VIRTUAL Identificacion Insaturacion y Grupo AlcoholDocumento9 páginasLABORATORIO VIRTUAL Identificacion Insaturacion y Grupo Alcoholdayis robayoAún no hay calificaciones
- II Examen Sumativo CEPUNT 2018-I (A)Documento17 páginasII Examen Sumativo CEPUNT 2018-I (A)Kevin Minchola Castañeda100% (2)
- Agua y MineralesDocumento11 páginasAgua y MineralesJuanjoSerranoAún no hay calificaciones
- Catálogo Muebles Oct20Documento56 páginasCatálogo Muebles Oct20wellasur001Aún no hay calificaciones
- La Bomba - MateoFlechaDocumento18 páginasLa Bomba - MateoFlechaClemente HernandezAún no hay calificaciones
- Manual Química OrgánicaDocumento56 páginasManual Química OrgánicaFermínAún no hay calificaciones
- Ficha Técnica Operaciones para La Creación y Producción de Una SillaDocumento2 páginasFicha Técnica Operaciones para La Creación y Producción de Una SillaSherley DuarteAún no hay calificaciones
- Practica 1 Metales PDFDocumento10 páginasPractica 1 Metales PDFMiguelAngelo music-y-gameplayAún no hay calificaciones
- Balance de MateriaDocumento6 páginasBalance de MateriaCROMO-ZINCADOS VALMOS S.A.SAún no hay calificaciones
- Preliminar Escalera QuimicaDocumento4 páginasPreliminar Escalera QuimicaedwinsazzzAún no hay calificaciones
- Carbón ActivadoDocumento6 páginasCarbón ActivadoSaul Marroquin HernandezAún no hay calificaciones
- Tarea 2Documento3 páginasTarea 2Matías MarambioAún no hay calificaciones
- Guia Once Funciones Oxigenadas 2 ParteDocumento24 páginasGuia Once Funciones Oxigenadas 2 Partesergio alarconAún no hay calificaciones
- Composicion de La Leche de Vaca - Cabra-Oveja para Quesos WordDocumento8 páginasComposicion de La Leche de Vaca - Cabra-Oveja para Quesos WordleonardoagudeloAún no hay calificaciones
- Ejercicios de Termodinámica I: Física y Química 1º BachilleratoDocumento2 páginasEjercicios de Termodinámica I: Física y Química 1º BachilleratoCesarHernandezAún no hay calificaciones
- Prueba Curso ExcelDocumento17 páginasPrueba Curso ExcelJoan HoyosAún no hay calificaciones
- Soluciones IiiDocumento2 páginasSoluciones IiiLuz Angela Perlas100% (1)
- Infraestructura de Un laboratorio-KMLDocumento3 páginasInfraestructura de Un laboratorio-KMLKari DominguezAún no hay calificaciones
- Titulación Potenciometrica y ConductimetricaDocumento18 páginasTitulación Potenciometrica y ConductimetricaAntonio Rivero100% (1)
- Sesión 2-19 de SeptiembreDocumento33 páginasSesión 2-19 de SeptiembreNeysis Ivania GomezAún no hay calificaciones
- Q SSM Dir Sem05 PDFDocumento2 páginasQ SSM Dir Sem05 PDFStiven AlvarezAún no hay calificaciones
- Plantas en Ambientes SalinosDocumento31 páginasPlantas en Ambientes SalinosSELENE VANESSA CHOZO MESTANZA100% (1)
- Proyecto Final - GeologiaDocumento9 páginasProyecto Final - GeologiacristianAún no hay calificaciones
- Mapa Conceptual - Rocas ÍgneasDocumento2 páginasMapa Conceptual - Rocas ÍgneasDeyner Ortiz QuispeAún no hay calificaciones
- Hoja de Seguridad - P1Documento3 páginasHoja de Seguridad - P1Axel Morales Piñón.Aún no hay calificaciones
- Coalescente TX Alveg TexanolDocumento11 páginasCoalescente TX Alveg Texanolkarla villagomezAún no hay calificaciones
- Lectura AutótrofosDocumento2 páginasLectura AutótrofosYaz PerezAún no hay calificaciones
- Conocer Los Diferentes Métodos de Esterilización Por Agentes Físicos y Químicos. Diferenciar Esterilización Entre Calor y Calor HumadoDocumento9 páginasConocer Los Diferentes Métodos de Esterilización Por Agentes Físicos y Químicos. Diferenciar Esterilización Entre Calor y Calor HumadoClinton LunaAún no hay calificaciones
- Guia de Estudio para El Taller Teorico Practico Sobre Disoluciones 4to AñoDocumento45 páginasGuia de Estudio para El Taller Teorico Practico Sobre Disoluciones 4to Añonando fernandoAún no hay calificaciones
- CABALLERO RAMIREZ Jose Omar - TemaDocumento9 páginasCABALLERO RAMIREZ Jose Omar - TemaChristian Collazos PardaveAún no hay calificaciones