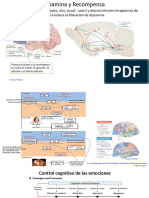
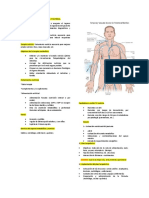
También podría gustarte
- Metabolismo del sistema digestivo, del hígado, de la vesícula y de las vías biliares: En condiciones de salud y en las enfermedadesDe EverandMetabolismo del sistema digestivo, del hígado, de la vesícula y de las vías biliares: En condiciones de salud y en las enfermedadesAún no hay calificaciones
- ¿Puede el cannabis ayudarte a perder peso?De Everand¿Puede el cannabis ayudarte a perder peso?Aún no hay calificaciones
- Bloque 2 - Lectura BaseDocumento66 páginasBloque 2 - Lectura BaseSebastian MorenoAún no hay calificaciones
- Mecanismos Regulatorios de La IngestiónDocumento6 páginasMecanismos Regulatorios de La IngestiónLihoy KamAún no hay calificaciones
- Bases Fisiopatológicas de La ObesidadDocumento7 páginasBases Fisiopatológicas de La ObesidadHalcon Negro DEL SurAún no hay calificaciones
- Regulacion Neuro-Endocrina y Control de Peso PDFDocumento58 páginasRegulacion Neuro-Endocrina y Control de Peso PDFCarlos García IbáñezAún no hay calificaciones
- Regulación de La IngestaDocumento3 páginasRegulación de La IngestaMaggie JacksonAún no hay calificaciones
- Mecanismos Regulatorios de La Ingestión de Alimentos - Navira Tello-NutricionDocumento7 páginasMecanismos Regulatorios de La Ingestión de Alimentos - Navira Tello-NutricionNavira TelloAún no hay calificaciones
- Hambre y SaciedadDocumento43 páginasHambre y SaciedadJaunleo100% (1)
- Ingestión de AlimentosDocumento12 páginasIngestión de AlimentosAlejandro Londoño AmarilesAún no hay calificaciones
- Fisiologia Del ApetitoDocumento3 páginasFisiologia Del Apetitojohanesquivelca0% (1)
- Mecanismos de RecompensaDocumento28 páginasMecanismos de RecompensaGaboGonzalezAún no hay calificaciones
- Fisiologia Hormonal IntestinalDocumento14 páginasFisiologia Hormonal IntestinalChristian Benjo Lopez EidAún no hay calificaciones
- Fisiología Hambre y SaciedadDocumento19 páginasFisiología Hambre y SaciedadDionisio RodríguezAún no hay calificaciones
- Papel Del NpyDocumento20 páginasPapel Del NpyAlonso Pérez PeraltaAún no hay calificaciones
- Conferencia Nutricion y Equilibrio EnergeticoDocumento29 páginasConferencia Nutricion y Equilibrio EnergeticoJacob NavarroAún no hay calificaciones
- Hambre y SaciedadDocumento35 páginasHambre y SaciedadMar OlaAún no hay calificaciones
- Fisiopatología de La ObesidadDocumento21 páginasFisiopatología de La ObesidadVictoria Diaz EstradaAún no hay calificaciones
- Interrelaciones Metabolicas UpaoDocumento32 páginasInterrelaciones Metabolicas UpaoAAaab100% (1)
- 7 Regulación de La Temperatura - LJFRDocumento48 páginas7 Regulación de La Temperatura - LJFRmmarquinas1Aún no hay calificaciones
- Marco Teorico 1Documento29 páginasMarco Teorico 1Mario Perez RumboAún no hay calificaciones
- GrelinaDocumento12 páginasGrelinaAdriana Cabrera ToledoAún no hay calificaciones
- 03 - La Ciencia de La ObesidadDocumento8 páginas03 - La Ciencia de La ObesidadElizabeth SotoAún no hay calificaciones
- Participación Del Sistema Nervioso y Del Tracto GastrointestinalDocumento11 páginasParticipación Del Sistema Nervioso y Del Tracto GastrointestinalFranklin Jose Rojas ColónAún no hay calificaciones
- Polipeptido PancreaticoDocumento2 páginasPolipeptido Pancreaticomanuel_vca100% (1)
- Obesidad y Resistencia A La Leptina PDFDocumento6 páginasObesidad y Resistencia A La Leptina PDFMaria CelesteAún no hay calificaciones
- Obesidad 19Documento43 páginasObesidad 1911-MH-HU-XIOMARA INGRID JAZMIN MARTINEZ ROMANIAún no hay calificaciones
- Enn U1 Ea KatmDocumento6 páginasEnn U1 Ea KatmKaty tm100% (1)
- Hambre y SedDocumento6 páginasHambre y SedPablo PazmiñoAún no hay calificaciones
- Regulación A Largo Plazo Del HambreDocumento7 páginasRegulación A Largo Plazo Del HambremariaaAún no hay calificaciones
- Trastornos de La AlimentacionDocumento58 páginasTrastornos de La AlimentacionMariana Argumedo SawczuckAún no hay calificaciones
- Gordos y FlacosDocumento27 páginasGordos y FlacosTati Del MarAún no hay calificaciones
- OBESIDADDocumento24 páginasOBESIDADMinerva García G.Aún no hay calificaciones
- Ayudantía 05Documento17 páginasAyudantía 05Claudia PizarroAún no hay calificaciones
- Encefalopatía PDFDocumento9 páginasEncefalopatía PDFAnto RomeroAún no hay calificaciones
- Determinantes Fisiológicos y Culturales de La Conducta AlimentariaDocumento9 páginasDeterminantes Fisiológicos y Culturales de La Conducta AlimentariaLeila RAún no hay calificaciones
- Conducta Al Comer Estudia4Documento8 páginasConducta Al Comer Estudia4marisol castilloAún no hay calificaciones
- Regulación Central Del MetabolismoDocumento4 páginasRegulación Central Del Metabolismogrupo6 mktAún no hay calificaciones
- Clase 6 ObesidadDocumento9 páginasClase 6 ObesidadCamila Rivas DíazAún no hay calificaciones
- Ud8 Tema 1 Bioquímica Del Control Del Apetito Guía de Actividades 2020Documento8 páginasUd8 Tema 1 Bioquímica Del Control Del Apetito Guía de Actividades 2020centrodecopiadotokio libreriaAún no hay calificaciones
- UntitledDocumento7 páginasUntitledBatsheba AsayagAún no hay calificaciones
- Ane223e UnlockedDocumento9 páginasAne223e UnlockedLuismi DocAún no hay calificaciones
- Factores Orexigénicos y Ambientales DefinitivoDocumento6 páginasFactores Orexigénicos y Ambientales DefinitivoAndreaAún no hay calificaciones
- Gamberale, M - Regulación de La Conducta AlimentariaDocumento14 páginasGamberale, M - Regulación de La Conducta AlimentariaKarim AlvarezAún no hay calificaciones
- Teoria M1T4 Regulacion Del ApetitoDocumento9 páginasTeoria M1T4 Regulacion Del ApetitoGonzalo MartinezAún no hay calificaciones
- REGULADORESNEUROENDOCRINOSYGASTROINTESTINALESDELAPETITOYLASACIEDADDocumento28 páginasREGULADORESNEUROENDOCRINOSYGASTROINTESTINALESDELAPETITOYLASACIEDADBarbaraAún no hay calificaciones
- DownloadDocumento12 páginasDownloadmaloga1985Aún no hay calificaciones
- Paciente CríticoDocumento52 páginasPaciente Críticoconstanza Aravena Faúndez100% (1)
- Neurofisiología Del Sistema Hambre SaciedadDocumento11 páginasNeurofisiología Del Sistema Hambre Saciedadapi-3854803Aún no hay calificaciones
- Terapia Metabólica y NutriciaDocumento26 páginasTerapia Metabólica y NutriciaAndy LinaresAún no hay calificaciones
- 2022 Diplomado NFyobesidad LipotrópicosDocumento42 páginas2022 Diplomado NFyobesidad LipotrópicoszayrumAún no hay calificaciones
- Afi2 U1 Ea MarzDocumento8 páginasAfi2 U1 Ea Marzmarisol100% (1)
- Ramírez José Cuadro Sinóptico Fisiología Saciedad Hambre PDFDocumento5 páginasRamírez José Cuadro Sinóptico Fisiología Saciedad Hambre PDFKyria DevilspardaAún no hay calificaciones
- Clase 9 Conducta Alimentaria y Regulacion Metabolica HormonalDocumento37 páginasClase 9 Conducta Alimentaria y Regulacion Metabolica HormonalLuís Ponte Ayala100% (1)
- M2-T4 Regulación Fisiológica de La AlimentaciónDocumento10 páginasM2-T4 Regulación Fisiológica de La AlimentaciónSil SerranoAún no hay calificaciones
- Hormonas GastronitesDocumento14 páginasHormonas GastronitesAbril VelásquezAún no hay calificaciones
- Obesidad Exógena FinalDocumento24 páginasObesidad Exógena FinalAna FloresAún no hay calificaciones
- YANOWSKYDocumento11 páginasYANOWSKYNely MendiotaAún no hay calificaciones
- 0000001137cnt 2017 09 Cuadernillo ObesidadDocumento36 páginas0000001137cnt 2017 09 Cuadernillo ObesidadcanalespabloAún no hay calificaciones
- Dieta Keto. ¿Saludable o peligrosa? Descubra riesgos ocultos según los medicos!De EverandDieta Keto. ¿Saludable o peligrosa? Descubra riesgos ocultos según los medicos!Aún no hay calificaciones
- Guia de Problemas 1 - 2021 CORREGIDADocumento6 páginasGuia de Problemas 1 - 2021 CORREGIDAFranco FigueroaAún no hay calificaciones
- Mafis 35 (1) 39-48Documento10 páginasMafis 35 (1) 39-48Franco FigueroaAún no hay calificaciones
- Trabajo Práctico #4: Pardeamiento Enzimático Y No EnzimáticoDocumento7 páginasTrabajo Práctico #4: Pardeamiento Enzimático Y No EnzimáticoFranco FigueroaAún no hay calificaciones
- Hormonas de La SaciedadDocumento6 páginasHormonas de La SaciedadFranco FigueroaAún no hay calificaciones
- Estudios Hidrologico Puente CbbaDocumento143 páginasEstudios Hidrologico Puente CbbaJhilmar AlcocerAún no hay calificaciones
- Analis de MicroempresariosDocumento8 páginasAnalis de MicroempresariosSHONA TUCHIA PETSAAún no hay calificaciones
- Ingles-Iiee2017-U4-Sesion 34Documento6 páginasIngles-Iiee2017-U4-Sesion 34Manuel Atarama Bayona100% (1)
- ManzanillaDocumento1 páginaManzanillaEsteban CruzAún no hay calificaciones
- Mecanica de FluidosDocumento16 páginasMecanica de FluidosCesar MenesesAún no hay calificaciones
- Guia 1.ciencias Naturales Bióticos y Abióticos. Grado SegundoDocumento3 páginasGuia 1.ciencias Naturales Bióticos y Abióticos. Grado SegundoLeonardo Velasco100% (1)
- Segundo ActoDocumento5 páginasSegundo ActoAndreaVilarAún no hay calificaciones
- Anatomia, Fisiología y SemiologiaDocumento6 páginasAnatomia, Fisiología y SemiologiaHugo CuevaAún no hay calificaciones
- Calma Tu SedDocumento5 páginasCalma Tu Sedfernando gonzalezAún no hay calificaciones
- Articulo de Aceite A Base de Semilla de MaracuyaDocumento13 páginasArticulo de Aceite A Base de Semilla de MaracuyaJunior CanchilaAún no hay calificaciones
- Historia Natural Del AcvDocumento22 páginasHistoria Natural Del Acvamordavid922280100% (1)
- Desarrollo Motor, Habilidades y Destrezas Del IndividuoDocumento12 páginasDesarrollo Motor, Habilidades y Destrezas Del IndividuoMaria RiveoAún no hay calificaciones
- Literatura 5 Secuencia Didáctica Sobre Comprensión LectoraDocumento4 páginasLiteratura 5 Secuencia Didáctica Sobre Comprensión LectoraIrenemmAún no hay calificaciones
- El Texto-Tipos de Textos (2024)Documento3 páginasEl Texto-Tipos de Textos (2024)amaliamadrid2211Aún no hay calificaciones
- Virus HerpesDocumento14 páginasVirus HerpesHernández Rayón Celeste AbigailAún no hay calificaciones
- CombinepdfDocumento11 páginasCombinepdfJuan RomainaAún no hay calificaciones
- Protocolo de ViolenciaDocumento24 páginasProtocolo de ViolenciaJesús Israel Nieves RomeroAún no hay calificaciones
- Luruaco Es Un Municipio ColombianoDocumento38 páginasLuruaco Es Un Municipio ColombianoMari ArroyoAún no hay calificaciones
- Cadenas TróficasDocumento4 páginasCadenas TróficasLeonardo PichardoAún no hay calificaciones
- De Musica de Pseudo PlutarcoDocumento3 páginasDe Musica de Pseudo PlutarcoRodrigoAún no hay calificaciones
- Informe de Quimica 2 Laboratorio # 4Documento21 páginasInforme de Quimica 2 Laboratorio # 4alfredoAún no hay calificaciones
- Taller de Sensibilización Fitness y SaludDocumento7 páginasTaller de Sensibilización Fitness y SaludCarlos Hoyos EspitiaAún no hay calificaciones
- Dos ValijasDocumento2 páginasDos ValijasBrenda RiviereAún no hay calificaciones
- Teoria Del SoporteDocumento28 páginasTeoria Del SoporteARTURO PICON MONTOYAAún no hay calificaciones
- Claves de Respuesta Cuestionario CEIDocumento4 páginasClaves de Respuesta Cuestionario CEISantiago Ortegón RestrepoAún no hay calificaciones
- Argumentacion JuridicaDocumento18 páginasArgumentacion JuridicaHBEnkiAún no hay calificaciones
- Inventario Fauna-Sierra de PenjamoDocumento12 páginasInventario Fauna-Sierra de PenjamoEmily CanoAún no hay calificaciones
- AporofobiaDocumento5 páginasAporofobiaalfmotattoAún no hay calificaciones
- Como Tratar Con Personas DificilesDocumento36 páginasComo Tratar Con Personas DificilesFerchi WerAún no hay calificaciones
- Carrión Feijoo Cinthia Thalia - PrácticaNº9 PDFDocumento3 páginasCarrión Feijoo Cinthia Thalia - PrácticaNº9 PDFCinthia CarrionAún no hay calificaciones