También podría gustarte
- Dr. Sebi Enfermedades Pulmonares Hierbas y Dietas Alcalinas: Detener Enfermedades o Inflamaciones Pulmonares, Tos, Asma, Tuberculosis, etc., y Eliminar las Mucosidades Infectadas del Tubo DigestivoDe EverandDr. Sebi Enfermedades Pulmonares Hierbas y Dietas Alcalinas: Detener Enfermedades o Inflamaciones Pulmonares, Tos, Asma, Tuberculosis, etc., y Eliminar las Mucosidades Infectadas del Tubo DigestivoAún no hay calificaciones
- Ventilación PulmonarDocumento53 páginasVentilación PulmonarAurora Gonzalez Lezcano100% (1)
- Conceptos Y Estrategias En Terapia RespiratoriaDe EverandConceptos Y Estrategias En Terapia RespiratoriaCalificación: 4 de 5 estrellas4/5 (1)
- Liberacion Flash y DiferencialDocumento2 páginasLiberacion Flash y DiferencialvanessaAún no hay calificaciones
- Fisiología Pulmonar 1Documento23 páginasFisiología Pulmonar 1ISABELLA DURAN CARDEL50% (2)
- Practica N8 QODocumento8 páginasPractica N8 QOCARRAQ9Aún no hay calificaciones
- Tipos de Núclidos y Especies Iónicas para Segundo Grado de SecundariaDocumento3 páginasTipos de Núclidos y Especies Iónicas para Segundo Grado de SecundariaRonald PalominoAún no hay calificaciones
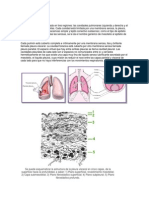
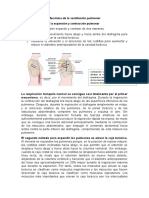
- Ventilación pulmonar: procesos y factoresDocumento22 páginasVentilación pulmonar: procesos y factoresYini Geovanelli0% (1)
- Fisiologia PulmonarDocumento8 páginasFisiologia PulmonarAnthony Aguilar PizánAún no hay calificaciones
- Sistema Respiratorio-Fisiología PDFDocumento16 páginasSistema Respiratorio-Fisiología PDFMaría José CabreraAún no hay calificaciones
- Fisiología RonielDocumento8 páginasFisiología RonielRoniel RoaAún no hay calificaciones
- Resumen Sistema RespiratorioDocumento15 páginasResumen Sistema RespiratoriomariaAún no hay calificaciones
- UNIDAD 6 SISTEMA RESPIRATORIODocumento24 páginasUNIDAD 6 SISTEMA RESPIRATORIOMicaela ArnaoAún no hay calificaciones
- Mecánica de La Ventilación PulmonarDocumento11 páginasMecánica de La Ventilación PulmonarJorge MoscosoAún no hay calificaciones
- Fisiologia Respiratoria IDocumento7 páginasFisiologia Respiratoria IReyes TobiasAún no hay calificaciones
- Fisiologia ResppDocumento6 páginasFisiologia ResppPatriciaJimenezAún no hay calificaciones
- CAVIDAD PLEURAL y Mecanismo de La VentilacionDocumento4 páginasCAVIDAD PLEURAL y Mecanismo de La VentilacionJhon Anthony Escudero Mejia100% (1)
- Ciclo RespiratorioDocumento4 páginasCiclo RespiratorioNutrición UIWAún no hay calificaciones
- Ventilacion Pulmonal 2Documento11 páginasVentilacion Pulmonal 2Jhobsen JesusAún no hay calificaciones
- TÉCNICA E INTERPRETACIÓN DE LA ESPIROMETRÍADocumento11 páginasTÉCNICA E INTERPRETACIÓN DE LA ESPIROMETRÍAAbrahan HaufAún no hay calificaciones
- Capitulo 38 Ventilación Pulmonar Jorge Palma 201942192Documento4 páginasCapitulo 38 Ventilación Pulmonar Jorge Palma 201942192Vivi Montenegro0% (1)
- UABP 4. Sistema RespiratorioDocumento29 páginasUABP 4. Sistema RespiratorioMartín PanicoAún no hay calificaciones
- Documento de HistologiaDocumento19 páginasDocumento de HistologiaEmily Gaibor EscobarAún no hay calificaciones
- Tema 7 Sistema RespiratorioDocumento8 páginasTema 7 Sistema RespiratorioMaria TorresAún no hay calificaciones
- Fisiología Respiratoria - Ventilación PulmonarDocumento15 páginasFisiología Respiratoria - Ventilación PulmonarNinoska Martinez100% (1)
- Funciones Del Sistema RespiratorioDocumento18 páginasFunciones Del Sistema RespiratorioEvelin LuxAún no hay calificaciones
- Biofisica de La Respiraciòn, 2014.Documento67 páginasBiofisica de La Respiraciòn, 2014.maria nAún no hay calificaciones
- Ventilacion Pulmonar PDFDocumento13 páginasVentilacion Pulmonar PDFLisbeth Alejandra Martínez GonzálezAún no hay calificaciones
- Taller de Fisiologia Del Sitema RespiratorioDocumento20 páginasTaller de Fisiologia Del Sitema RespiratorioYeimy Caraballo Patiño100% (1)
- Conceptos Básicos de Mecánica VentilatoriaDocumento6 páginasConceptos Básicos de Mecánica VentilatoriaDyma Osorio NaviaAún no hay calificaciones
- Presiones RespiratoriasDocumento8 páginasPresiones RespiratoriasMayté Chuchuca MiteAún no hay calificaciones
- Respiracion en La AlturaDocumento43 páginasRespiracion en La AlturaRobert Berrocal QuispeAún no hay calificaciones
- 4 Mecanica RespiratoriaDocumento16 páginas4 Mecanica Respiratoriajira1987Aún no hay calificaciones
- Reporte de Lectura Capítulo 38 Ventilación PulmonarDocumento10 páginasReporte de Lectura Capítulo 38 Ventilación PulmonarMiguel CórdovaAún no hay calificaciones
- Mecanica RespiratoriaDocumento5 páginasMecanica RespiratoriaKatheryn Lezama GuerraAún no hay calificaciones
- ApuntesDocumento7 páginasApuntesAbel VillalbaAún no hay calificaciones
- Clase 6 Fisiologia - Ventilacion PulmonarDocumento5 páginasClase 6 Fisiologia - Ventilacion PulmonarJaqui De La CruzAún no hay calificaciones
- Resumen Sobre Los Mecanismos de La Respiración PulmonarDocumento6 páginasResumen Sobre Los Mecanismos de La Respiración PulmonarJunior ArenasAún no hay calificaciones
- NEUMOLOGIADocumento6 páginasNEUMOLOGIAFreddy SupeAún no hay calificaciones
- Resumen de RespiracionDocumento11 páginasResumen de RespiracionPame NuniAún no hay calificaciones
- Revisión anatomía y fisiología respiratoriaDocumento14 páginasRevisión anatomía y fisiología respiratoriagiorealAún no hay calificaciones
- Ventilacion PulmunarDocumento26 páginasVentilacion Pulmunarleonardoalfonso.garcia25Aún no hay calificaciones
- Músculos pulmonaresDocumento14 páginasMúsculos pulmonaresAnaAún no hay calificaciones
- Monografia Fisiologia Del EjercicioDocumento6 páginasMonografia Fisiologia Del EjercicioHellen Mallaupoma ObregonAún no hay calificaciones
- Ventilación Pulmonar y Función RespiratoriaDocumento4 páginasVentilación Pulmonar y Función RespiratoriaAlvaro Sergio SanchezAún no hay calificaciones
- Marco TeoricoDocumento8 páginasMarco TeoricoMateoAún no hay calificaciones
- Generalidades Del Sistema RespiratorioDocumento10 páginasGeneralidades Del Sistema RespiratorioMartina BergerAún no hay calificaciones
- Sist RespiratorioDocumento20 páginasSist RespiratorioJose RomeroAún no hay calificaciones
- Anatomia y Fisiologia de Las VrespiratoriasDocumento2 páginasAnatomia y Fisiologia de Las Vrespiratoriasnathaliecastillo758Aún no hay calificaciones
- Unidad 2. Ventilación PulmonarDocumento43 páginasUnidad 2. Ventilación PulmonarMaria Elena OrtizAún no hay calificaciones
- MecánicaRespiratoria.J8Documento11 páginasMecánicaRespiratoria.J8Alisson BeitiaAún no hay calificaciones
- Conferencia Función RespiratoriaDocumento14 páginasConferencia Función RespiratoriaJerselita RuizAún no hay calificaciones
- Clase 2-Fisiologia Respiratoria IiDocumento6 páginasClase 2-Fisiologia Respiratoria IiGina BatshebaAún no hay calificaciones
- MFH IV RespiratorioDocumento51 páginasMFH IV RespiratorioAmara BestAún no hay calificaciones
- Fisiologia RespiratoriaDocumento51 páginasFisiologia RespiratoriaMarco Pereira MaysAún no hay calificaciones
- Sistema RespiratorioDocumento16 páginasSistema RespiratorioJoyas Cosmeticos SolangeAún no hay calificaciones
- Fisiologia RespiratoriaDocumento14 páginasFisiologia RespiratoriaSTEPHANIE PAOLA VAZQUEZ ORTEGAAún no hay calificaciones
- Ventilación Pulmonar PDFDocumento6 páginasVentilación Pulmonar PDFJeffrey LoayzaAún no hay calificaciones
- Mecánica VentilatoriaDocumento6 páginasMecánica VentilatoriaSofia GarciaAún no hay calificaciones
- BCT MusculosDocumento58 páginasBCT MusculosAlejandra Tenorio HernándezAún no hay calificaciones
- Resumen Del Resumen FisiologíaDocumento14 páginasResumen Del Resumen FisiologíacamAún no hay calificaciones
- RESPIRATORIO I y CARDIACO FISIOLOGIADocumento31 páginasRESPIRATORIO I y CARDIACO FISIOLOGIAJuu MoranAún no hay calificaciones
- Tema 4Documento19 páginasTema 4Irene CulpeperAún no hay calificaciones
- 10-PPT-Estado GaseosoDocumento14 páginas10-PPT-Estado GaseosoJulio Soto LovonAún no hay calificaciones
- Taller de La Semana 5 (V)Documento11 páginasTaller de La Semana 5 (V)Joseph Jahuira TaparaAún no hay calificaciones
- TermodinámicaDocumento3 páginasTermodinámicaWendy CantorAún no hay calificaciones
- Cristalización azúcar UNAN LeónDocumento11 páginasCristalización azúcar UNAN Leónsidar espinozaAún no hay calificaciones
- Redox IiDocumento11 páginasRedox IiAlejandra Mendez MansillaAún no hay calificaciones
- Método ConductimétricoDocumento5 páginasMétodo Conductimétrico708170100% (2)
- FISICA Ciclo TermodinamicoDocumento1 páginaFISICA Ciclo TermodinamicoAngel SalamancaAún no hay calificaciones
- Pruebas de Caracterización de Derivados HalogenadosDocumento11 páginasPruebas de Caracterización de Derivados HalogenadosIsabella MariñoAún no hay calificaciones
- Lab No.3 Metodo VolumetricoDocumento7 páginasLab No.3 Metodo VolumetricoDelia GentoAún no hay calificaciones
- Previo 6 Prueba ADocumento3 páginasPrevio 6 Prueba AAlejandro Garcia GarciaAún no hay calificaciones
- Actividad 3.1 - Balances de Energía Con Reacción QuímicaDocumento4 páginasActividad 3.1 - Balances de Energía Con Reacción QuímicaJocelyn Grisel García GonzálezAún no hay calificaciones
- Tension Superficial de Un LiquidoDocumento7 páginasTension Superficial de Un Liquidojunior jimenezAún no hay calificaciones
- Átomo y Sus CaracterísticasDocumento2 páginasÁtomo y Sus CaracterísticasSusana Lorena Ospina MaldonadoAún no hay calificaciones
- Informe Diseño Operacional de Una Torre de Destilación - Acetona-CloroformoDocumento12 páginasInforme Diseño Operacional de Una Torre de Destilación - Acetona-CloroformoESTEFANIA VELEZ GUZMANAún no hay calificaciones
- TEMA 3 CALOR Y TRABAJO MECANICO V2 Santa CruzDocumento70 páginasTEMA 3 CALOR Y TRABAJO MECANICO V2 Santa CruzFabiana CuellarAún no hay calificaciones
- Ejercicios de Bioquímica sobre Cinética Enzimática de Michaelis-Menten y Lineweaver-BurkDocumento2 páginasEjercicios de Bioquímica sobre Cinética Enzimática de Michaelis-Menten y Lineweaver-Burkfernando maldonadoAún no hay calificaciones
- Tema 4Documento19 páginasTema 4Loreto BlancoAún no hay calificaciones
- Resina AcrílicaDocumento16 páginasResina AcrílicaElizabeth ArizonaAún no hay calificaciones
- Especificaciones Materias PrimasDocumento26 páginasEspecificaciones Materias Primasjesy2012Aún no hay calificaciones
- Fuerzas intermoleculares: tipos y ejemplosDocumento11 páginasFuerzas intermoleculares: tipos y ejemplosJohan SanherAún no hay calificaciones
- Reservorios IIDocumento67 páginasReservorios IIMarcosMarinAún no hay calificaciones
- Ejercicios Resueltos - SERIE I - 1ley de Termodinámica PDFDocumento24 páginasEjercicios Resueltos - SERIE I - 1ley de Termodinámica PDFJuan Pablo IngrassiaAún no hay calificaciones
- Guia#2 Propiedades TermodinamicasDocumento3 páginasGuia#2 Propiedades Termodinamicasjeremy gabrielAún no hay calificaciones
- Ejercicios Resueltos Módulo 4-2Documento11 páginasEjercicios Resueltos Módulo 4-2Rodrigo CastroAún no hay calificaciones
- Taller Semana 7Documento4 páginasTaller Semana 7jhassury jhammely pocomucha chuquillanqui0% (1)
- Asignación 5Documento4 páginasAsignación 5Grecia Cajigas VelásquezAún no hay calificaciones
- Soxhlet 2Documento18 páginasSoxhlet 2gamaAún no hay calificaciones