También podría gustarte
- Conceptos Y Estrategias En Terapia RespiratoriaDe EverandConceptos Y Estrategias En Terapia RespiratoriaCalificación: 4 de 5 estrellas4/5 (1)
- Inhala y exhala: Conoce tus pulmonesDe EverandInhala y exhala: Conoce tus pulmonesCalificación: 3.5 de 5 estrellas3.5/5 (3)
- Ciclo RespiratorioDocumento4 páginasCiclo RespiratorioNutrición UIWAún no hay calificaciones
- Fisiologia Respiratoria IDocumento7 páginasFisiologia Respiratoria IReyes TobiasAún no hay calificaciones
- Tema 29 Fisiologia PropioDocumento11 páginasTema 29 Fisiologia PropiojoseluAún no hay calificaciones
- Anatomía RespiratoriaDocumento3 páginasAnatomía RespiratoriaErika RodriguezRodriguezAún no hay calificaciones
- Temas Respiratorio Fisiologia PDFDocumento32 páginasTemas Respiratorio Fisiologia PDFjoseluAún no hay calificaciones
- Sistema Respiratorio-Fisiología PDFDocumento16 páginasSistema Respiratorio-Fisiología PDFMaría José CabreraAún no hay calificaciones
- Sist Respiratorio 1Documento3 páginasSist Respiratorio 1Maria Florencia LopezAún no hay calificaciones
- Fisiologia PulmonarDocumento8 páginasFisiologia PulmonarAnthony Aguilar PizánAún no hay calificaciones
- 023respiratorio (2020)Documento20 páginas023respiratorio (2020)Bonitas de MariaAún no hay calificaciones
- RespiratorioDocumento3 páginasRespiratoriocarolina ordoñezAún no hay calificaciones
- Resumen Sistema RespiratorioDocumento15 páginasResumen Sistema RespiratoriomariaAún no hay calificaciones
- Sistema Respiratorio (Marco Teórico)Documento5 páginasSistema Respiratorio (Marco Teórico)Liolaemus Tenuis100% (6)
- Unidad 6 Sistema RespiratorioDocumento24 páginasUnidad 6 Sistema RespiratorioMicaela ArnaoAún no hay calificaciones
- Taller de Fisiologia Del Sitema RespiratorioDocumento20 páginasTaller de Fisiologia Del Sitema RespiratorioYeimy Caraballo Patiño100% (1)
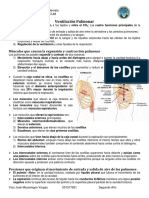
- Ventilación PulmonarDocumento22 páginasVentilación PulmonarYini Geovanelli0% (1)
- Respuestas y Adaptaciones Pulmonares Al EjercicioDocumento47 páginasRespuestas y Adaptaciones Pulmonares Al EjercicioYusmari CarielAún no hay calificaciones
- Conceptos Básicos de Mecánica VentilatoriaDocumento6 páginasConceptos Básicos de Mecánica VentilatoriaDyma Osorio NaviaAún no hay calificaciones
- Sistema RespiratorioDocumento16 páginasSistema RespiratorioJoyas Cosmeticos SolangeAún no hay calificaciones
- Sistema RespiratorioDocumento9 páginasSistema RespiratorioroostellAún no hay calificaciones
- Taller Sistema Respiratorio MetodologiaDocumento6 páginasTaller Sistema Respiratorio MetodologiaGenerith BolañoAún no hay calificaciones
- Clase Biologia Sistema RespiratorioDocumento18 páginasClase Biologia Sistema RespiratorioFacundoAún no hay calificaciones
- Resumen de RespiracionDocumento11 páginasResumen de RespiracionPame NuniAún no hay calificaciones
- 3.3. Sistema RespiratorioDocumento9 páginas3.3. Sistema RespiratorioBereniceAún no hay calificaciones
- Fisiologia RespiratoriaDocumento45 páginasFisiologia RespiratoriaraphaelAún no hay calificaciones
- Taller de Respiracioon Plantas y Animales PDFDocumento5 páginasTaller de Respiracioon Plantas y Animales PDFAlejandro Alfonzo Ruiz Lopez100% (1)
- VentilaciónDocumento9 páginasVentilaciónLisset Cifuentes MirandaAún no hay calificaciones
- Mecánica de La Ventilación PulmonarDocumento11 páginasMecánica de La Ventilación PulmonarJorge MoscosoAún no hay calificaciones
- Fisiología Respiratoria Guia - 20240222 - 081950 - 0000Documento8 páginasFisiología Respiratoria Guia - 20240222 - 081950 - 0000Jesús ReyesAún no hay calificaciones
- Resumen - Fisiologia RespiratoriaDocumento16 páginasResumen - Fisiologia RespiratoriaAndres MunozAún no hay calificaciones
- Mecánica RespiratoriaDocumento20 páginasMecánica RespiratoriaMoises VindasAún no hay calificaciones
- Apuntes Del Sistema RespiratorioDocumento14 páginasApuntes Del Sistema RespiratorioAdriana López-ValverdeAún no hay calificaciones
- Fisiologia RespiratoriaDocumento5 páginasFisiologia RespiratoriaFuentes SoniaAún no hay calificaciones
- La Fisiología de La RespiraciónDocumento7 páginasLa Fisiología de La RespiraciónLeonardo CabreraAún no hay calificaciones
- UABP 4. Sistema RespiratorioDocumento29 páginasUABP 4. Sistema RespiratorioMartín PanicoAún no hay calificaciones
- Capitulo 38 Ventilación Pulmonar Jorge Palma 201942192Documento4 páginasCapitulo 38 Ventilación Pulmonar Jorge Palma 201942192Vivi Montenegro0% (1)
- Anatomia Fisiologia Del Aparato RespiratorioDocumento13 páginasAnatomia Fisiologia Del Aparato RespiratorioJohn Steve Auquilla AlaoAún no hay calificaciones
- UNIDAD II: Cuidado de Las Personas Con Alteraciones en La Necesidad de Oxigenación.Documento6 páginasUNIDAD II: Cuidado de Las Personas Con Alteraciones en La Necesidad de Oxigenación.Daiana RojasAún no hay calificaciones
- Sist RespiratorioDocumento20 páginasSist RespiratorioJose RomeroAún no hay calificaciones
- Af1-Mod 1-RespDocumento12 páginasAf1-Mod 1-RespGustavo CamaraAún no hay calificaciones
- Respiracion en La AlturaDocumento43 páginasRespiracion en La AlturaRobert Berrocal QuispeAún no hay calificaciones
- Fisiologia Respiratoria HumanaDocumento27 páginasFisiologia Respiratoria Humanayuglennys amayaAún no hay calificaciones
- Mecánica VentilatoriaDocumento6 páginasMecánica VentilatoriaSofia GarciaAún no hay calificaciones
- Biofisica Respiratoria 2022-1Documento56 páginasBiofisica Respiratoria 2022-1Daniel CabarcasAún no hay calificaciones
- RespiraciónDocumento110 páginasRespiraciónMore ParedesAún no hay calificaciones
- Ensayo Semana 1Documento4 páginasEnsayo Semana 1mouAún no hay calificaciones
- Sistema Respiratorio FisioDocumento84 páginasSistema Respiratorio FisioMaria José Narro GarciaAún no hay calificaciones
- FisiologiaDocumento6 páginasFisiologiaNurlind RachidAún no hay calificaciones
- RespiracionDocumento6 páginasRespiracionCarmen BonillaAún no hay calificaciones
- La Respiración Es Un Proceso FísicoDocumento3 páginasLa Respiración Es Un Proceso FísicoMirame Otra Vez100% (3)
- Resumen Aparato Respiratorio - Fisio.Documento8 páginasResumen Aparato Respiratorio - Fisio.Giannina GattiAún no hay calificaciones
- Biofisica Mecanica Respiratoria IDocumento128 páginasBiofisica Mecanica Respiratoria IFrank Simbaña MoncadaAún no hay calificaciones
- Guia Octavos Sistema RespiratorioDocumento3 páginasGuia Octavos Sistema RespiratorioAly AravenaAún no hay calificaciones
- Fisiología Respiratoria PDFDocumento3 páginasFisiología Respiratoria PDFYasna VAún no hay calificaciones
- Fisiología PulmonarDocumento67 páginasFisiología PulmonarGrecia Paulina Padilla100% (1)
- Trabajo Práctico #14 Mecanica Ventilatoria (M.A)Documento9 páginasTrabajo Práctico #14 Mecanica Ventilatoria (M.A)Martina ArneraAún no hay calificaciones
- Fisiología Respiratoria ResumenDocumento5 páginasFisiología Respiratoria Resumenarmando guitierrezAún no hay calificaciones
- Cap 37, Ventilación Pulmonar, GuytonDocumento10 páginasCap 37, Ventilación Pulmonar, GuytonLizzy GrillAún no hay calificaciones
- Fisiología Respiratoria - Ventilación PulmonarDocumento15 páginasFisiología Respiratoria - Ventilación PulmonarNinoska Martinez100% (1)
- 3.4 DPRDocumento3 páginas3.4 DPRAlex PerezAún no hay calificaciones
- 06 Examen Enero Sexto Grado 2020Documento11 páginas06 Examen Enero Sexto Grado 2020Cosme VelascoAún no hay calificaciones
- Evaluación Diagnóstica CTA - 2°Documento7 páginasEvaluación Diagnóstica CTA - 2°Miriam Calderon SalvatierraAún no hay calificaciones
- Aparto DigestivoDocumento19 páginasAparto Digestivoeleazar_2002481Aún no hay calificaciones
- Manual Tgu Por Jeronimo Milo 1Documento45 páginasManual Tgu Por Jeronimo Milo 1Mauricio CarvajalAún no hay calificaciones
- Ficha Técnica Geomalla UniaxialDocumento9 páginasFicha Técnica Geomalla UniaxialGonzalo Guerra VegaAún no hay calificaciones
- SexualidadDocumento3 páginasSexualidadMaría Fernanda AguilarAún no hay calificaciones
- Horario Clases 1-2023Documento1 páginaHorario Clases 1-2023RAUL ELICEO ZAMBRANA OCAÑAAún no hay calificaciones
- BTL Sis Presentacion EspañolDocumento75 páginasBTL Sis Presentacion EspañolMatt ShowAún no hay calificaciones
- Quiroga, Horacio - La Miel SilvestreDocumento4 páginasQuiroga, Horacio - La Miel SilvestreAdrian BarahonaAún no hay calificaciones
- Apuntes Sobre MicroorganimosDocumento13 páginasApuntes Sobre MicroorganimosclauAún no hay calificaciones
- Calificación de Un Área Aséptica de Uso Docencia en La Planta Piloto Farmacéutica de La Fes Zaragoza UnamDocumento14 páginasCalificación de Un Área Aséptica de Uso Docencia en La Planta Piloto Farmacéutica de La Fes Zaragoza Unammmartinezr26095836Aún no hay calificaciones
- Impacto Diferencial Del Estrés Entre Hombres y MujeresDocumento16 páginasImpacto Diferencial Del Estrés Entre Hombres y MujeresHanna BethAún no hay calificaciones
- Neolítico UrbanoDocumento5 páginasNeolítico UrbanoHector Luis Parra GonzalezAún no hay calificaciones
- System Description - FlexiROC T D C 50-65 Actualizada 2017 PDFDocumento161 páginasSystem Description - FlexiROC T D C 50-65 Actualizada 2017 PDFErick Jonathan Nuñez Bugueño100% (3)
- Soltero, Casado, Viudo y DivorciadoDocumento33 páginasSoltero, Casado, Viudo y DivorciadoCarlos A Medina FAún no hay calificaciones
- Informe #4. Superficies Equipotenciales y Líneas de Campo eléctrico.M2BDocumento6 páginasInforme #4. Superficies Equipotenciales y Líneas de Campo eléctrico.M2BArteta Alvarez Daniel EstebanAún no hay calificaciones
- Planimetría CintaDocumento32 páginasPlanimetría CintaAledj Martinez100% (1)
- 72 Nombres de DiosDocumento13 páginas72 Nombres de DiosAmeliaesmeraldaAún no hay calificaciones
- El Derecho Socializado - Uladech 2017Documento24 páginasEl Derecho Socializado - Uladech 2017ROSITAAún no hay calificaciones
- Biorremediacion 1Documento1 páginaBiorremediacion 1Katherine Rocio Obeid ManjarrezAún no hay calificaciones
- Correción Parcial GeneticaDocumento20 páginasCorreción Parcial GeneticaAnonymous EPm5OyGAún no hay calificaciones
- Intriago - Sistemas Ópticos Inalámbricos en El Espectro Visible (VLC) para Comunicaciones e Ilumi...Documento53 páginasIntriago - Sistemas Ópticos Inalámbricos en El Espectro Visible (VLC) para Comunicaciones e Ilumi...adheMarceloAún no hay calificaciones
- UNIDAD 1. Clasificación de La Materia - Ejercicios PropuestosDocumento2 páginasUNIDAD 1. Clasificación de La Materia - Ejercicios Propuestosalexis ortegaAún no hay calificaciones
- DONOFRIODocumento12 páginasDONOFRIOJanetAún no hay calificaciones
- CORRELACIóN ESTRATIGRAFICA - OKDocumento15 páginasCORRELACIóN ESTRATIGRAFICA - OKJhonatan Valenzuela H100% (1)
- Agua y Desague para Ciudad ConstitucionDocumento138 páginasAgua y Desague para Ciudad ConstitucionJhonAbelAún no hay calificaciones
- Herramientas Ingenieria IndustrialDocumento8 páginasHerramientas Ingenieria IndustrialScm PaqueteriaAún no hay calificaciones
- Los 7 Desperdicios de La Construcción - La Construcción Sin PérdidasDocumento2 páginasLos 7 Desperdicios de La Construcción - La Construcción Sin PérdidasMartha MezarinoAún no hay calificaciones
- Movimiento de TierraDocumento62 páginasMovimiento de TierraRoderick Xavier CastilloAún no hay calificaciones