También podría gustarte
- Aportes de la biología del suelo a la agroecologíaDe EverandAportes de la biología del suelo a la agroecologíaCalificación: 5 de 5 estrellas5/5 (1)
- Identificación de hongos asociados a suelos agrícolasDe EverandIdentificación de hongos asociados a suelos agrícolasAún no hay calificaciones
- Taller Metodo Matricial de VigasDocumento10 páginasTaller Metodo Matricial de VigasJesus MoralesAún no hay calificaciones
- Turismo GastronómicoDocumento21 páginasTurismo GastronómicoPERLA YONARA NAZARENO ANGULOAún no hay calificaciones
- Fósforo, Potasio y AzufreDocumento21 páginasFósforo, Potasio y AzufreDelfina MaderoAún no hay calificaciones
- Trabajo de CalculoDocumento59 páginasTrabajo de CalculoFredy Octavio CHAPARRO LUNAAún no hay calificaciones
- Universidad Nacional Agraria de La SelvaDocumento13 páginasUniversidad Nacional Agraria de La SelvaLeninEguiLingánArellanoAún no hay calificaciones
- El Ciclo Del PotacioDocumento8 páginasEl Ciclo Del PotacioEdpa Duran MaciasAún no hay calificaciones
- Dinámica Del FósforoDocumento3 páginasDinámica Del FósforoJean Carlos DelgadoAún no hay calificaciones
- Balance de NutrientesDocumento7 páginasBalance de NutrientesXimena RAAún no hay calificaciones
- Balance de FosforoDocumento7 páginasBalance de FosforoHernán Castro SanabriaAún no hay calificaciones
- Fertilizantes QuímicosDocumento3 páginasFertilizantes QuímicosGothardo Casanova TorresAún no hay calificaciones
- Microbios mediadores de cambios en la vegetación por fertilización N y PDocumento15 páginasMicrobios mediadores de cambios en la vegetación por fertilización N y Pmirian escalante oreAún no hay calificaciones
- Abono Organico de La Lombriz San RafaelDocumento56 páginasAbono Organico de La Lombriz San RafaelDaysy ParedesAún no hay calificaciones
- Traducción Articulo SolubilizaciónDocumento6 páginasTraducción Articulo SolubilizaciónHury GaldámezAún no hay calificaciones
- 01 - Fertilidad Del SueloDocumento15 páginas01 - Fertilidad Del SueloCesar Max Perlacios VargasAún no hay calificaciones
- La importancia de la materia orgánica en los suelosDocumento5 páginasLa importancia de la materia orgánica en los suelosAndy MuñozAún no hay calificaciones
- Traducido IIDocumento8 páginasTraducido IIBelsy AlfaroAún no hay calificaciones
- Enmiendas Agricolas - OrganicasDocumento5 páginasEnmiendas Agricolas - OrganicasDaniel MontalvoAún no hay calificaciones
- Relación C / N en Los SuelosDocumento10 páginasRelación C / N en Los SuelosGustavoArgoteAún no hay calificaciones
- Biomasa Microbiana Del SueloDocumento8 páginasBiomasa Microbiana Del SueloGonzalo RestrepoAún no hay calificaciones
- 41 1 74-91 Osorio - Microorganismos Del Suelo y Disponibilidad de NutrientesDocumento18 páginas41 1 74-91 Osorio - Microorganismos Del Suelo y Disponibilidad de Nutrientesjuank_1983Aún no hay calificaciones
- Química Aplicada 3Documento12 páginasQuímica Aplicada 3Fabiana GautoAún no hay calificaciones
- Taller 5 Analisis Articulo Biofertilizantes Alternativa Biotecnologica para Los AgroecosistemasDocumento5 páginasTaller 5 Analisis Articulo Biofertilizantes Alternativa Biotecnologica para Los AgroecosistemasMelissa BenitesAún no hay calificaciones
- NR40100Documento21 páginasNR40100Bastian SeguelAún no hay calificaciones
- Tarea5 - Grupo - 7Documento31 páginasTarea5 - Grupo - 7natalia ramirezAún no hay calificaciones
- Trabajo de Lechuga.Documento19 páginasTrabajo de Lechuga.Rossii Acosta Ortiz0% (1)
- Caso 5 Fijacion de FosforoDocumento9 páginasCaso 5 Fijacion de FosforoFERNANDOAún no hay calificaciones
- ABONAMIENTO Y RESIEMBRA DE PASTOS Y FORRAJES LeoDocumento6 páginasABONAMIENTO Y RESIEMBRA DE PASTOS Y FORRAJES LeoLilian SalazarAún no hay calificaciones
- Diferentes Ciclos de Los NutrientesDocumento10 páginasDiferentes Ciclos de Los NutrientesBLANCA ROSA ALVAREZ LOPEZAún no hay calificaciones
- Informe de Materia Organica Del SueloDocumento11 páginasInforme de Materia Organica Del SueloDiaz AndresAún no hay calificaciones
- ResumenDocumento5 páginasResumenjose armando fabian salvadorAún no hay calificaciones
- La Disponibilidad Del Fósforo Es Producto de La Actividad Bacteriana en El Suelo en Ecosistemas Oligotróficos: Una Revisión CríticaDocumento12 páginasLa Disponibilidad Del Fósforo Es Producto de La Actividad Bacteriana en El Suelo en Ecosistemas Oligotróficos: Una Revisión Críticajorge silvaAún no hay calificaciones
- ASPECTOS BÁSICOS DEL MANEJO DEL FÓSFORO - Carlos PutulDocumento25 páginasASPECTOS BÁSICOS DEL MANEJO DEL FÓSFORO - Carlos PutulOsias CaalAún no hay calificaciones
- Tesis Grado.Documento20 páginasTesis Grado.Franco ydkdaAún no hay calificaciones
- Biofertilizantes aumentan cosecha caña azúcarDocumento15 páginasBiofertilizantes aumentan cosecha caña azúcarDiego FdoAún no hay calificaciones
- Py Nutrientes Del Suelo y La Dendidad de Raíz en AguajeDocumento35 páginasPy Nutrientes Del Suelo y La Dendidad de Raíz en AguajeOscar CastroAún no hay calificaciones
- Interacción Fósforo - ZincDocumento28 páginasInteracción Fósforo - ZincFerxu MontoyaAún no hay calificaciones
- 95-Texto Del Artículo-236-1-10-20180424Documento14 páginas95-Texto Del Artículo-236-1-10-20180424Laura Semeraro MongeAún no hay calificaciones
- Informe de Caracterización Bioquímica de Microorganismos Rizosféricos de Plantas de Vainilla Con Potencial Como BiofertilizantesDocumento4 páginasInforme de Caracterización Bioquímica de Microorganismos Rizosféricos de Plantas de Vainilla Con Potencial Como BiofertilizantesWILLIAM EDUARDO PICOAún no hay calificaciones
- BOCASHI Generalidades de Los Abonos OrgánicosDocumento12 páginasBOCASHI Generalidades de Los Abonos OrgánicosAbrahamAún no hay calificaciones
- Sintesis MODocumento2 páginasSintesis MOGriselda LopezAún no hay calificaciones
- Introducción Suelo, AbonoDocumento8 páginasIntroducción Suelo, AbonoRaúl Solís PérezAún no hay calificaciones
- Materia orgánica sueloDocumento3 páginasMateria orgánica sueloAdrian LuceroAún no hay calificaciones
- Respuestas Fisiológicas de Plantas Cultivadas en Bioespacios Por Efecto de Nanofertilizantes y ZeolitaDocumento18 páginasRespuestas Fisiológicas de Plantas Cultivadas en Bioespacios Por Efecto de Nanofertilizantes y ZeolitaMáximo GuillénAún no hay calificaciones
- Cap 9 y 10 SintesisDocumento5 páginasCap 9 y 10 Sintesisricardo.ramosc11Aún no hay calificaciones
- Materia Organica 3.1Documento4 páginasMateria Organica 3.1Nikole MoralesAún no hay calificaciones
- Indicadores Biológicos y Funciones Del SueloDocumento7 páginasIndicadores Biológicos y Funciones Del SueloVictoria RodriguezAún no hay calificaciones
- Manejo Sostenible de SuelosDocumento15 páginasManejo Sostenible de Suelosluis angel soto palacinAún no hay calificaciones
- Materia orgánica suelo efectosDocumento3 páginasMateria orgánica suelo efectosAndrés TrujilloAún no hay calificaciones
- Materia OrganicaDocumento4 páginasMateria OrganicaPPLUCHOAún no hay calificaciones
- Suelitos Materia Orgánica Marco TeoricoDocumento9 páginasSuelitos Materia Orgánica Marco TeoricoLuciaWarryAún no hay calificaciones
- CLASE 3. Fijacion de Nitrogeno, Disponibilidad y Extracción de Nutrientes de Los Pastos y Forrajes. Programa de Fertilización EstratégicaDocumento54 páginasCLASE 3. Fijacion de Nitrogeno, Disponibilidad y Extracción de Nutrientes de Los Pastos y Forrajes. Programa de Fertilización Estratégicaelio carboAún no hay calificaciones
- Ecologia MonografiaDocumento8 páginasEcologia MonografiaRon RomeroAún no hay calificaciones
- Uso de Composta y Humus de LombrizDocumento7 páginasUso de Composta y Humus de LombrizGabriela Hernandez SuarezAún no hay calificaciones
- Microorganismos Solubilizadores de Fosforo FinalDocumento18 páginasMicroorganismos Solubilizadores de Fosforo FinalLADY ANGELICA NAULA GONZALEZAún no hay calificaciones
- El PH Del Suelo en La AgriculturaDocumento4 páginasEl PH Del Suelo en La AgriculturaCristian Andres Grast JuradoAún no hay calificaciones
- Manejo Del Fósforo en Maíz Tesis 1Documento4 páginasManejo Del Fósforo en Maíz Tesis 1Javier Guzman HernandezAún no hay calificaciones
- Materia Organica Del SueloDocumento5 páginasMateria Organica Del SueloOscar Palomino PomaAún no hay calificaciones
- Biochar como enmienda agrícola: efectos sobre propiedades del suelo y rasgos funcionales de las plantasDocumento4 páginasBiochar como enmienda agrícola: efectos sobre propiedades del suelo y rasgos funcionales de las plantaspedrocqAún no hay calificaciones
- Microbios del suelo y su papel en la agriculturaDocumento21 páginasMicrobios del suelo y su papel en la agriculturaERBIN ROJAS50% (2)
- Efecto Del Potasio A Diferentes Concentraciones y Compuestos, en El MaizDocumento28 páginasEfecto Del Potasio A Diferentes Concentraciones y Compuestos, en El MaizVeronica BermudoAún no hay calificaciones
- Evaluación Del Fertilizante Orgánico Líquido de Lombriz San RafaelDocumento11 páginasEvaluación Del Fertilizante Orgánico Líquido de Lombriz San RafaelUziel MartinezAún no hay calificaciones
- INGLES Avalancha de Nuevas Motos para Colombia en La Feria 2 Ruedas 2022Documento6 páginasINGLES Avalancha de Nuevas Motos para Colombia en La Feria 2 Ruedas 2022Daivid Sebastian Muñoz RoballoAún no hay calificaciones
- Informe Inventario Forestal Del Barrio Senderos de Santa MartaDocumento18 páginasInforme Inventario Forestal Del Barrio Senderos de Santa MartaDaivid Sebastian Muñoz RoballoAún no hay calificaciones
- Sistema Silvoagricola GranjaDocumento33 páginasSistema Silvoagricola GranjaDaivid Sebastian Muñoz RoballoAún no hay calificaciones
- Formulario Registro Fotografico 1Documento10 páginasFormulario Registro Fotografico 1Daivid Sebastian Muñoz RoballoAún no hay calificaciones
- Precios Marfil 16 MarzoDocumento6 páginasPrecios Marfil 16 MarzoDaivid Sebastian Muñoz RoballoAún no hay calificaciones
- Capitulo 1Documento8 páginasCapitulo 1Daivid Sebastian Muñoz RoballoAún no hay calificaciones
- Informe Del DocumentoDocumento10 páginasInforme Del DocumentoDaivid Sebastian Muñoz RoballoAún no hay calificaciones
- Bioluminescence Style Minitheme by SlidesgoDocumento10 páginasBioluminescence Style Minitheme by SlidesgoDaivid Sebastian Muñoz RoballoAún no hay calificaciones
- Abstracta ArtDocumento24 páginasAbstracta ArtDaivid Sebastian Muñoz RoballoAún no hay calificaciones
- Proyecto de AulaDocumento3 páginasProyecto de AulaDaivid Sebastian Muñoz RoballoAún no hay calificaciones
- El Papel de Los Microorganismos Del Suelo enDocumento14 páginasEl Papel de Los Microorganismos Del Suelo enDaivid Sebastian Muñoz RoballoAún no hay calificaciones
- Preguntas Del ExamenDocumento8 páginasPreguntas Del ExamenDaivid Sebastian Muñoz RoballoAún no hay calificaciones
- Cientifico MicrobiologiaDocumento2 páginasCientifico MicrobiologiaDaivid Sebastian Muñoz RoballoAún no hay calificaciones
- Capitulo 3Documento8 páginasCapitulo 3Daivid Sebastian Muñoz RoballoAún no hay calificaciones
- Capitulo 2Documento9 páginasCapitulo 2Daivid Sebastian Muñoz RoballoAún no hay calificaciones
- Clase Procesal Laboral 05 de Abril de 2022Documento39 páginasClase Procesal Laboral 05 de Abril de 2022Daivid Sebastian Muñoz RoballoAún no hay calificaciones
- Cientifico MicrobiologiaDocumento2 páginasCientifico MicrobiologiaDaivid Sebastian Muñoz RoballoAún no hay calificaciones
- Estado de Resultado: Fecha Tipo Vendedor DescripciónDocumento4 páginasEstado de Resultado: Fecha Tipo Vendedor DescripciónDaivid Sebastian Muñoz RoballoAún no hay calificaciones
- Formatos Fotos SebastianDocumento1 páginaFormatos Fotos SebastianDaivid Sebastian Muñoz RoballoAún no hay calificaciones
- Preguntas Del Parcial Final Procesal LaboralDocumento7 páginasPreguntas Del Parcial Final Procesal LaboralDaivid Sebastian Muñoz RoballoAún no hay calificaciones
- Resumen Del Video de BioeticaDocumento7 páginasResumen Del Video de BioeticaDaivid Sebastian Muñoz RoballoAún no hay calificaciones
- Propuesta de ValorDocumento1 páginaPropuesta de ValorDaivid Sebastian Muñoz RoballoAún no hay calificaciones
- Derecho Laboral HistoriaDocumento16 páginasDerecho Laboral HistoriaDaivid Sebastian Muñoz RoballoAún no hay calificaciones
- Presupuesto AsdrubalDocumento136 páginasPresupuesto AsdrubalDaivid Sebastian Muñoz RoballoAún no hay calificaciones
- Sistema ProductivoDocumento17 páginasSistema ProductivoDaivid Sebastian Muñoz RoballoAún no hay calificaciones
- Resumen Del Video de BioeticaDocumento7 páginasResumen Del Video de BioeticaDaivid Sebastian Muñoz RoballoAún no hay calificaciones
- Avance Proyecto DiseñoDocumento14 páginasAvance Proyecto DiseñoDaivid Sebastian Muñoz RoballoAún no hay calificaciones
- Resumen Del Video de BioeticaDocumento7 páginasResumen Del Video de BioeticaDaivid Sebastian Muñoz RoballoAún no hay calificaciones
- Resumen Del Video de BioeticaDocumento7 páginasResumen Del Video de BioeticaDaivid Sebastian Muñoz RoballoAún no hay calificaciones
- PASOS PARA CALCULO DE UNA LOSA mACIZA EN 1 DIRECCIONDocumento5 páginasPASOS PARA CALCULO DE UNA LOSA mACIZA EN 1 DIRECCIONHarrison JesúsAún no hay calificaciones
- Asis Departamental Cundinamarca 2022Documento263 páginasAsis Departamental Cundinamarca 2022María Ximena Gómez SuárezAún no hay calificaciones
- Anexo 2Documento2 páginasAnexo 2Justa Carmen Concha RevillaAún no hay calificaciones
- Practica Radiación de Cuerpo Negro Con Simulador (Física Cuántica)Documento3 páginasPractica Radiación de Cuerpo Negro Con Simulador (Física Cuántica)Fátima QuijasAún no hay calificaciones
- Seminario-5.6 14-IiDocumento2 páginasSeminario-5.6 14-IiGracce KellyAún no hay calificaciones
- Optimizar operaciones bodegasDocumento19 páginasOptimizar operaciones bodegasMichael GonzalezAún no hay calificaciones
- Tales de MiletoDocumento5 páginasTales de MiletoAcount FeikAún no hay calificaciones
- Centrifuga y Centripeta, Estatica, Facultad de Ingenieria, UNAMDocumento3 páginasCentrifuga y Centripeta, Estatica, Facultad de Ingenieria, UNAMEduardo Daniel Batta GonzalezAún no hay calificaciones
- Lo Fundamental de Todo Proceso Pedagógico Es El Aprendizaje y NoDocumento6 páginasLo Fundamental de Todo Proceso Pedagógico Es El Aprendizaje y NolizziAún no hay calificaciones
- Diagrama de Flujo Del CaféDocumento9 páginasDiagrama de Flujo Del CaféAlexCajasAún no hay calificaciones
- Modulo 3 Kelly SamillanDocumento7 páginasModulo 3 Kelly SamillankellyAún no hay calificaciones
- Vco R4 PDFDocumento6 páginasVco R4 PDFAlex Diaz VargasAún no hay calificaciones
- Maquinaria para La Elaboración de EmbutidosDocumento9 páginasMaquinaria para La Elaboración de EmbutidosPérez Yac María MercedesAún no hay calificaciones
- Guia 1. Determinación de Humedad y Solidos Totales en AlimentosDocumento2 páginasGuia 1. Determinación de Humedad y Solidos Totales en AlimentosDiana CristinaAún no hay calificaciones
- El Mago de Los ColoresDocumento4 páginasEl Mago de Los ColoresGaby musicAún no hay calificaciones
- Operar Camión Minero de Acuerdo Con Manual Operativo y Normativas TécnicasDocumento5 páginasOperar Camión Minero de Acuerdo Con Manual Operativo y Normativas Técnicasing.alfredoherrera.a8494Aún no hay calificaciones
- Noticias AlimarketDocumento13 páginasNoticias AlimarketCcoo MahouAún no hay calificaciones
- Cuadernillo La Letra DDocumento13 páginasCuadernillo La Letra DJeanette TrincadoAún no hay calificaciones
- Ebook - Estrategias para Mejorar El Clima LaboralDocumento7 páginasEbook - Estrategias para Mejorar El Clima LaboralServicios y Distribuidora JM c.aAún no hay calificaciones
- Tipos de Hernias ClasfDocumento3 páginasTipos de Hernias ClasfJosé HernándezAún no hay calificaciones
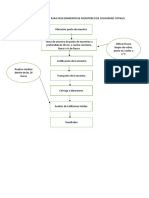
- Diagrama de Bloque para Procedimiento de Monitoreo de Coliformes TotalesDocumento3 páginasDiagrama de Bloque para Procedimiento de Monitoreo de Coliformes TotalesRoberto BrunaAún no hay calificaciones
- Manual Del Maestro: Aldo Lavagnini (Magister)Documento108 páginasManual Del Maestro: Aldo Lavagnini (Magister)jnunez59Aún no hay calificaciones
- FILOSOFÍA Trabajo ExposiciónDocumento31 páginasFILOSOFÍA Trabajo ExposicióncrisAún no hay calificaciones
- Parashat Bo: Resumen de La Parashá Comentario A La ParasháDocumento3 páginasParashat Bo: Resumen de La Parashá Comentario A La ParasháisraelAún no hay calificaciones
- Nematodos NematomorfosDocumento9 páginasNematodos NematomorfosLuis Adrian Gomez ArceAún no hay calificaciones
- Reino Moneras, Hongos y ProtoctistaDocumento7 páginasReino Moneras, Hongos y ProtoctistaCarolay BrievaAún no hay calificaciones
- Agente Reductor NopalDocumento4 páginasAgente Reductor NopalJivi' Gernot DunckelAún no hay calificaciones