También podría gustarte
- FOTOSINTESISDocumento3 páginasFOTOSINTESIShnnc100% (44)
- Plantas C3, C4 Y CAMDocumento8 páginasPlantas C3, C4 Y CAMAlejandro Cueto100% (1)
- Plantas c3, c4 y CamDocumento11 páginasPlantas c3, c4 y CamRony PérezAún no hay calificaciones
- FOTOSINTESISDocumento23 páginasFOTOSINTESISVivian Martínez PiérolaAún no hay calificaciones
- 3 IUPAC Libro Rojo 2005 EspañolDocumento382 páginas3 IUPAC Libro Rojo 2005 EspañolCamilo Andres Rico Cortes100% (6)
- Linfoma de HodgkinDocumento20 páginasLinfoma de HodgkinMarianella Saravia LopezAún no hay calificaciones
- FotosintesisDocumento21 páginasFotosintesisJanexy NohelyAún no hay calificaciones
- Potencial industrial de los residuos de la caña de azúcarDocumento20 páginasPotencial industrial de los residuos de la caña de azúcarDavid Colorado VegaAún no hay calificaciones
- Acceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012De EverandAcceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012Aún no hay calificaciones
- Sintesis de La FotosintesisDocumento6 páginasSintesis de La FotosintesisAndrés GarnicaAún no hay calificaciones
- Tarea 4 Equipo 1Documento9 páginasTarea 4 Equipo 1brian antonio garciaAún no hay calificaciones
- Ciclo BensonDocumento5 páginasCiclo BensonMauri MtzAún no hay calificaciones
- ¿Qué Es La FotosíntesisDocumento2 páginas¿Qué Es La FotosíntesisJose BerganzaAún no hay calificaciones
- FotosintesisDocumento39 páginasFotosintesisalissonAún no hay calificaciones
- FotosintesisDocumento7 páginasFotosintesisfranco rosasAún no hay calificaciones
- Informe N°5 - FotosintesisDocumento19 páginasInforme N°5 - FotosintesisGianella Flor Carhuas PerezAún no hay calificaciones
- Parcial BiologiaDocumento4 páginasParcial BiologiaDANIELA ESTEFANNI CHAPARRO CHAPARROAún no hay calificaciones
- FotosíntesisDocumento3 páginasFotosíntesisDavid Aaron Diaz RicoAún no hay calificaciones
- Fotosintesis Eap Agroind2012Documento38 páginasFotosintesis Eap Agroind2012Rojas Mendoza Jesus100% (1)
- FotosintesisDocumento39 páginasFotosintesisKatty Cordova SalazarAún no hay calificaciones
- Universidad Autónoma AgrariaDocumento7 páginasUniversidad Autónoma Agrariaalfredomendieta459Aún no hay calificaciones
- Biologia Guia de La FotosintesisDocumento8 páginasBiologia Guia de La Fotosintesisgina celeste betancourtAún no hay calificaciones
- Ciclo de CalvinDocumento11 páginasCiclo de CalvinKriss Almendra Cieza QuevedoAún no hay calificaciones
- Reporte Practica 2Documento24 páginasReporte Practica 2Miguel QuevedoAún no hay calificaciones
- 7mo Teorico Anual SM 2023 ABC - PPTX ModificadoDocumento25 páginas7mo Teorico Anual SM 2023 ABC - PPTX ModificadoemersonespinovelasquezAún no hay calificaciones
- Tipos de fotosíntesis C3, C4 y CAMDocumento23 páginasTipos de fotosíntesis C3, C4 y CAMJULIO GUILLERMO CAHUANA CHUMBESAún no hay calificaciones
- Introducción A La FotosíntesisDocumento3 páginasIntroducción A La FotosíntesisAdrián Andrade RiveraAún no hay calificaciones
- Identificaciòn de Plantas C3 y C4Documento8 páginasIdentificaciòn de Plantas C3 y C4Jose Jacinto Magaña KaamalAún no hay calificaciones
- Importancia de La Fotosintesis en Los Pastos y ForrajesDocumento5 páginasImportancia de La Fotosintesis en Los Pastos y ForrajesPatricia ReKjoAún no hay calificaciones
- Tema 12 FotorrespiracionDocumento5 páginasTema 12 FotorrespiracionJose Natividad Flores MayoriAún no hay calificaciones
- Guia Sobre FotosintesisDocumento5 páginasGuia Sobre FotosintesisCarola LastraAún no hay calificaciones
- fotosintesisDocumento22 páginasfotosintesisIsabel González HernándezAún no hay calificaciones
- Punto 1 BCDocumento3 páginasPunto 1 BCfrancisco vieraAún no hay calificaciones
- Cadena Transportadora de ElectronesDocumento8 páginasCadena Transportadora de Electroneszulycastillo_2496Aún no hay calificaciones
- Cuál Es La Importancia de La HojaDocumento8 páginasCuál Es La Importancia de La HojaLu SanchezAún no hay calificaciones
- FOTOSINTESISDocumento11 páginasFOTOSINTESISDaniela De La Torre LeónAún no hay calificaciones
- Fotosintesis 175205 Downloable 1371027Documento4 páginasFotosintesis 175205 Downloable 1371027Joel UzuriagaAún no hay calificaciones
- Plantas C3 C4 CAM, PracticaDocumento5 páginasPlantas C3 C4 CAM, PracticaG Miller E EspírituAún no hay calificaciones
- Anatomia FoliarDocumento4 páginasAnatomia FoliarAngelica Margarito CanoAún no hay calificaciones
- ¿Qué Diferencia Hay Entre Plantas C3, C4 Y Cam?Documento9 páginas¿Qué Diferencia Hay Entre Plantas C3, C4 Y Cam?JUAN CAMILO FRANCO PENAAún no hay calificaciones
- Fotosintesis: Profesor Daniel Moena FlandesDocumento34 páginasFotosintesis: Profesor Daniel Moena FlandesAnarcos Andres Canivilo VargasAún no hay calificaciones
- Informe 2Documento5 páginasInforme 2Laura HerreraAún no hay calificaciones
- Reporte Práctica 2.Documento25 páginasReporte Práctica 2.José Daniel Jocop100% (1)
- Fotosintesis y La Respiracion Lab de BiologiaDocumento4 páginasFotosintesis y La Respiracion Lab de BiologiaDaniela Sánchez MedinaAún no hay calificaciones
- If 3 FotosintesisDocumento31 páginasIf 3 FotosintesisDenis PeraltaAún no hay calificaciones
- Fisio LoDocumento57 páginasFisio LoClaudia ReaAún no hay calificaciones
- 13 Anabolismo Foto y QuimiosintesisDocumento3 páginas13 Anabolismo Foto y Quimiosintesisbiopantera3875Aún no hay calificaciones
- Photosynthesis Science Presentation in Green Beige Illustrative StyleDocumento42 páginasPhotosynthesis Science Presentation in Green Beige Illustrative StyleAlejandro MG (Pana)Aún no hay calificaciones
- Mecanismos de concentración de CO2 en plantasDocumento87 páginasMecanismos de concentración de CO2 en plantasJHAMILKA ARHASY VILLEGAS CHEVEZAún no hay calificaciones
- FotosintesisDocumento2 páginasFotosintesisAna Cecilia EspinosaAún no hay calificaciones
- FotorrespiraciónDocumento2 páginasFotorrespiraciónCesar A ArangoAún no hay calificaciones
- Investigar "Fotosintesis en Plantas C4, Plantas Cam (Metabolismo Acido de Las Crasulaceas) y Fotorrespiracion"Documento15 páginasInvestigar "Fotosintesis en Plantas C4, Plantas Cam (Metabolismo Acido de Las Crasulaceas) y Fotorrespiracion"keisy DerasAún no hay calificaciones
- C3, C4, CamDocumento22 páginasC3, C4, CamMarisa LiziAún no hay calificaciones
- Cuestionario de FofotisntesisDocumento14 páginasCuestionario de FofotisntesisSANDRA NICOL MARQUINA ORBEGOZOAún no hay calificaciones
- ANÁLISIS DE RENDIMIENTO FOTOSINTÉTICO EN Elodea Sp.Documento11 páginasANÁLISIS DE RENDIMIENTO FOTOSINTÉTICO EN Elodea Sp.Dylan Alexander Rodriguez DiazAún no hay calificaciones
- Fijacion Del CarbonoDocumento9 páginasFijacion Del CarbonoCesar LópezAún no hay calificaciones
- Fase 3. Conocer El Proceso de Fotosíntesis y Metabolismo en Las PlantasDocumento14 páginasFase 3. Conocer El Proceso de Fotosíntesis y Metabolismo en Las PlantasJohan DiazAún no hay calificaciones
- CLOROPLASTOSDocumento11 páginasCLOROPLASTOSPaola RodríguezAún no hay calificaciones
- Fotosintesis RespiraciónDocumento28 páginasFotosintesis RespiraciónRosana ContrerasAún no hay calificaciones
- Fase Oscura de La FotosíntesisDocumento11 páginasFase Oscura de La FotosíntesisEDGAR1187Aún no hay calificaciones
- Guia teorica Eje 41 - Productores y fotosintesisDocumento8 páginasGuia teorica Eje 41 - Productores y fotosintesisconstanzagonzalez183Aún no hay calificaciones
- Química de macrocomponentes de alimentosDe EverandQuímica de macrocomponentes de alimentosAún no hay calificaciones
- Taller de Fisicoquimica-2Documento3 páginasTaller de Fisicoquimica-2Camilo Andres Rico CortesAún no hay calificaciones
- Punto 3Documento3 páginasPunto 3Camilo Andres Rico CortesAún no hay calificaciones
- Punto 5. Taller Fund. EspDocumento2 páginasPunto 5. Taller Fund. EspCamilo Andres Rico CortesAún no hay calificaciones
- Formato Informe FotosíntesisDocumento10 páginasFormato Informe FotosíntesisCamilo Andres Rico CortesAún no hay calificaciones
- Taller 1 EA-I-2021Documento9 páginasTaller 1 EA-I-2021Camilo Andres Rico CortesAún no hay calificaciones
- Seminario Grupo 5Documento13 páginasSeminario Grupo 5Camilo Andres Rico CortesAún no hay calificaciones
- pH soluciones amortiguadoras tallerDocumento2 páginaspH soluciones amortiguadoras tallerCamilo Andres Rico CortesAún no hay calificaciones
- Espectroscopia aplicada a compuestos orgánicos (2015572-1Documento4 páginasEspectroscopia aplicada a compuestos orgánicos (2015572-1Camilo Andres Rico CortesAún no hay calificaciones
- Informe ADNDocumento13 páginasInforme ADNCamilo Andres Rico CortesAún no hay calificaciones
- Informe I Aminoacidos Grupo 5Documento12 páginasInforme I Aminoacidos Grupo 5Camilo Andres Rico CortesAún no hay calificaciones
- Trabajo Miembros InferioresDocumento32 páginasTrabajo Miembros InferioresAngie Bernaez80% (5)
- Indice de Confort de La VegetacionDocumento22 páginasIndice de Confort de La VegetacionOrlando Jiménez GonzálezAún no hay calificaciones
- MetronidazolDocumento3 páginasMetronidazolSofia CastilloAún no hay calificaciones
- Introducción Al Sistema Respiratorio y VentilaciónDocumento14 páginasIntroducción Al Sistema Respiratorio y VentilaciónNelson RangelAún no hay calificaciones
- 5.tipos de PecesDocumento3 páginas5.tipos de Pecesana maria depaz barrionuevoAún no hay calificaciones
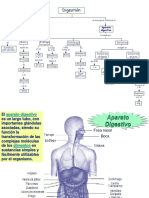
- Aparato DigestivoDocumento53 páginasAparato DigestivoRosario Paredes100% (1)
- CA 5 El Suelo Los Organismos Que Lo HabitanDocumento22 páginasCA 5 El Suelo Los Organismos Que Lo HabitanJohan Sebastián Chávez MosqueraAún no hay calificaciones
- Sistema EsqueléticoDocumento3 páginasSistema EsqueléticoSandra Jacqueline Peña ValdésAún no hay calificaciones
- Teorias de La EvoluciónDocumento3 páginasTeorias de La EvoluciónRUBEN DARIO MERCADO SALCEDOAún no hay calificaciones
- Soluciones Tampones y Sistemas Tampones Biológicos (San Juan Bautista)Documento14 páginasSoluciones Tampones y Sistemas Tampones Biológicos (San Juan Bautista)Renzo Alfredo Céspedes HerediaAún no hay calificaciones
- Agroecología y Transformaciones Culturales - Thomas León SicardDocumento25 páginasAgroecología y Transformaciones Culturales - Thomas León SicardLAURA INES CASTRO BARREROAún no hay calificaciones
- CUENTO - Un Hongo y Un Microbio Detras de Una AventuraDocumento1 páginaCUENTO - Un Hongo y Un Microbio Detras de Una AventuraFavio Mauricio GonzaAún no hay calificaciones
- Marco TeoricoDocumento4 páginasMarco TeoricoMariuxi Rocío Fajardo GuamanAún no hay calificaciones
- Relación Entre Delfines y Humanos PDFDocumento2 páginasRelación Entre Delfines y Humanos PDFpsvd19Aún no hay calificaciones
- Trabajo Nro 3Documento29 páginasTrabajo Nro 3Arven CastilloAún no hay calificaciones
- Adolf HitlerDocumento2 páginasAdolf HitlerMI La TinajaAún no hay calificaciones
- Tendinitis Del Supraespinoso PDFDocumento18 páginasTendinitis Del Supraespinoso PDFManuel Elescano100% (1)
- La Alucinación Por Ayahuasca (Jacques Mabit)Documento19 páginasLa Alucinación Por Ayahuasca (Jacques Mabit)drakpoAún no hay calificaciones
- Bi or Reactor EsDocumento11 páginasBi or Reactor EsGaby LissAún no hay calificaciones
- Bibliografia de Educacion AmbientalDocumento10 páginasBibliografia de Educacion Ambientaljose0% (1)
- 1 Qué Es El Estado de AprendizajeDocumento7 páginas1 Qué Es El Estado de AprendizajeGjgcAún no hay calificaciones
- Infografia 2 - 7 REPLICACION, TRANSCRIPCION Y TRADUCCION DEL ADNDocumento3 páginasInfografia 2 - 7 REPLICACION, TRANSCRIPCION Y TRADUCCION DEL ADNYERSON DANILO MORENO HUAROTOAún no hay calificaciones
- La expiación: celos y secretos entre amigosDocumento22 páginasLa expiación: celos y secretos entre amigosterefAún no hay calificaciones
- Agentes Tóxicos Generados Durante El Procesamiento de AlimentosDocumento3 páginasAgentes Tóxicos Generados Durante El Procesamiento de AlimentosGustavoAlexanderHernandezAún no hay calificaciones
- Caña de Azucar 2015Documento66 páginasCaña de Azucar 2015Frank Luque Alcarraz0% (1)
- Tema Vii-C0ntr0l Neural Del C0mp0rtamient0 AnimalDocumento14 páginasTema Vii-C0ntr0l Neural Del C0mp0rtamient0 AnimalDahibel HodgeAún no hay calificaciones
- Anatomía examen preguntasDocumento1143 páginasAnatomía examen preguntasDiego100% (2)
- S1 B UnjfscDocumento14 páginasS1 B UnjfscEddie Adrián Flores DíazAún no hay calificaciones