También podría gustarte
- El Origen de La Vida - Antonio LazcanoDocumento116 páginasEl Origen de La Vida - Antonio LazcanoAmoxcalli Ixcahuicopa88% (25)
- Radicales de Alcanos Alquenos y AlquinosDocumento7 páginasRadicales de Alcanos Alquenos y AlquinosElsie Esther0% (1)
- Artículo de Revista - Simbiosis y Evolución - Nicolás José Lavagnino, Alicia Massarini y Guillermo FolgueraDocumento21 páginasArtículo de Revista - Simbiosis y Evolución - Nicolás José Lavagnino, Alicia Massarini y Guillermo FolgueraFranco VillalbaAún no hay calificaciones
- La Teoría de La Evolución Como Revolución Conceptual Del MundoDocumento29 páginasLa Teoría de La Evolución Como Revolución Conceptual Del MundoMariano HorganAún no hay calificaciones
- 319028030002Documento19 páginas319028030002maria ferrerAún no hay calificaciones
- Documento Completo - PDF PDFADocumento11 páginasDocumento Completo - PDF PDFAAndy Quispe LuqueAún no hay calificaciones
- ANTROPOLOGÍA INTEGRAL - Marulanda PDFDocumento5 páginasANTROPOLOGÍA INTEGRAL - Marulanda PDFCatherine MarulandaAún no hay calificaciones
- Glosario de Terminos Grupalde La Autopoiesis Social Del Desarrollo Rural SustentableDocumento6 páginasGlosario de Terminos Grupalde La Autopoiesis Social Del Desarrollo Rural SustentableDunia Suarez BolivarAún no hay calificaciones
- SISTEMÁTICADocumento7 páginasSISTEMÁTICAeganchozo9668Aún no hay calificaciones
- Evolución en MéxicoDocumento9 páginasEvolución en MéxicoDeth GonzálezAún no hay calificaciones
- Interdisciplina y Falsas InterDocumento11 páginasInterdisciplina y Falsas InterMaria RestonAún no hay calificaciones
- BCNHumaniza REVISIONEu 4Documento25 páginasBCNHumaniza REVISIONEu 4EugeniaAún no hay calificaciones
- 17 Evolucion y Sus Repercusiones SocialesDocumento151 páginas17 Evolucion y Sus Repercusiones SocialesIniciativa Abierta AcAún no hay calificaciones
- Trabajo de AntropologiaDocumento13 páginasTrabajo de AntropologiaNaomichg GuevaraAún no hay calificaciones
- Partes y Funciones en El Desarrollo de La BiologiaDocumento10 páginasPartes y Funciones en El Desarrollo de La Biologiaisaac salcidoAún no hay calificaciones
- Emilio Muñoz Páginas 56-62Documento7 páginasEmilio Muñoz Páginas 56-62tiempos canallasAún no hay calificaciones
- La Síntesis Biosocial Una Propuesta Teórica en Antropología BiológicaDocumento13 páginasLa Síntesis Biosocial Una Propuesta Teórica en Antropología BiológicaClaudia CeronAún no hay calificaciones
- UNR - BIOANTROPOLOGÏA Ficha Unidades I, II, IIIDocumento11 páginasUNR - BIOANTROPOLOGÏA Ficha Unidades I, II, IIIBarbaAntropólogaAún no hay calificaciones
- 4 Biologia Filosofica Un Puente Epistemologico Entre La Biologia Mecanicista y Las Ciencias Humanosociales PDFDocumento16 páginas4 Biologia Filosofica Un Puente Epistemologico Entre La Biologia Mecanicista y Las Ciencias Humanosociales PDFbflorezg79Aún no hay calificaciones
- Taller Plan Lector Segunda Lengua Sofia MejiaDocumento4 páginasTaller Plan Lector Segunda Lengua Sofia MejiagrindrmateoriveraAún no hay calificaciones
- A4 Portada de Propuesta de Proyecto Científico Infantil Doodle Azul y NaranjaDocumento8 páginasA4 Portada de Propuesta de Proyecto Científico Infantil Doodle Azul y NaranjaMya QueenAún no hay calificaciones
- Ev 13725Documento19 páginasEv 13725Yeimi GonzalezAún no hay calificaciones
- Bio I Tema I Introduccion A La BiologiaDocumento5 páginasBio I Tema I Introduccion A La BiologiaxploratorAún no hay calificaciones
- Las Corrientes de Pensamiento Que Origen El Problema AmbientalDocumento6 páginasLas Corrientes de Pensamiento Que Origen El Problema Ambientalsanchezmarquez636Aún no hay calificaciones
- Cambio conceptual y representacional en el aprendizaje y la enseñanza de la cienciaDe EverandCambio conceptual y representacional en el aprendizaje y la enseñanza de la cienciaCalificación: 5 de 5 estrellas5/5 (1)
- Ensayo - Biodiversidad y Recuros Naturales-Marcos Alexis Romero BarónDocumento4 páginasEnsayo - Biodiversidad y Recuros Naturales-Marcos Alexis Romero BarónMarcos Alexis RomeroAún no hay calificaciones
- El Pensamiento Ambiental Como Génesis de La Bioética en La Investigación SocialDocumento10 páginasEl Pensamiento Ambiental Como Génesis de La Bioética en La Investigación SocialJOSE FABIAN RIOSAún no hay calificaciones
- Biologia Ensayo de El Libro Ecologia de PoblacionesDocumento4 páginasBiologia Ensayo de El Libro Ecologia de PoblacionesCrayolita MozZhAún no hay calificaciones
- Modelos para El Estudio Del DesarrolloDocumento10 páginasModelos para El Estudio Del DesarrolloStefania Coronel100% (1)
- Ensayo, SistematicaDocumento6 páginasEnsayo, SistematicaJHONATAN GARCIA100% (1)
- Resumen de Antro BIO 1Documento113 páginasResumen de Antro BIO 1Malvina HarispeAún no hay calificaciones
- Teoria Sintetica de La Evolucion2Documento6 páginasTeoria Sintetica de La Evolucion2Nahomi SantillanaAún no hay calificaciones
- Significado y Alcance de Pensar en Sistemas VivosDocumento33 páginasSignificado y Alcance de Pensar en Sistemas VivosOliver LudwingAún no hay calificaciones
- Implicaciones Epistemológicas Del Curriculum TransdisciplinarioDocumento28 páginasImplicaciones Epistemológicas Del Curriculum TransdisciplinarioLoedy Espinoza RaldezAún no hay calificaciones
- Andrade Ontogenia y FilogeniaDocumento35 páginasAndrade Ontogenia y FilogeniaJess Chávez MiguelAún no hay calificaciones
- Oyama Ken - Nuevos Paradigmas Y Fronteras en EcologiaDocumento0 páginasOyama Ken - Nuevos Paradigmas Y Fronteras en EcologiaEilyn VillegasAún no hay calificaciones
- Biosemiótica PDFDocumento22 páginasBiosemiótica PDFAlfonso Cárdenas PáezAún no hay calificaciones
- Ciclos VitalesDocumento12 páginasCiclos VitaleslicnoeliaAún no hay calificaciones
- TP 1 Antropologia Biologica UBA B MALOMODocumento7 páginasTP 1 Antropologia Biologica UBA B MALOMOBelu BatiahAún no hay calificaciones
- Desarrollo de La Antropologia Biologica en ColombiaDocumento28 páginasDesarrollo de La Antropologia Biologica en ColombiaCamilo Andres Contreras ValenciaAún no hay calificaciones
- La Teoría Biológica Del Conocimiento Como Puente de Articulación Entre Las Ciencias Naturales y SocialesDocumento12 páginasLa Teoría Biológica Del Conocimiento Como Puente de Articulación Entre Las Ciencias Naturales y SocialesMadre TierraAún no hay calificaciones
- Transdisciplinariedad Multi InterDocumento19 páginasTransdisciplinariedad Multi InterIliana SánchezAún no hay calificaciones
- Capítulo 3Documento23 páginasCapítulo 3alexandra carvajalarevaloAún no hay calificaciones
- El Cambio Epistemologico en Tres Funcionarios Publicos de La Cuarta TransformacionDocumento146 páginasEl Cambio Epistemologico en Tres Funcionarios Publicos de La Cuarta TransformacionRodriguez Barragán ElenaAún no hay calificaciones
- Biologia Evolucion Ecologia y Ambiente 3ero MedioDocumento77 páginasBiologia Evolucion Ecologia y Ambiente 3ero MedioJessica RiquelmeAún no hay calificaciones
- Aspectos Historicos y Epistemologicos Del Concepto de BiodiversidadDocumento10 páginasAspectos Historicos y Epistemologicos Del Concepto de BiodiversidadSergio Giovanny Gutierrez HernandezAún no hay calificaciones
- Dokumen - Tips - Anibal Rafael Abreu Santelises Id Ub08097ses14785 Nace La Comunicacion yDocumento15 páginasDokumen - Tips - Anibal Rafael Abreu Santelises Id Ub08097ses14785 Nace La Comunicacion yanibalabreuAún no hay calificaciones
- El Centauro OncologicoDocumento3 páginasEl Centauro OncologicoAndres PerillaAún no hay calificaciones
- Reseña EpistmologiaDocumento4 páginasReseña EpistmologiaJHONATAN VARGASAún no hay calificaciones
- Fundamentos Teóricos Del SonidoDocumento11 páginasFundamentos Teóricos Del Sonidoeric hernandezAún no hay calificaciones
- Ensayo 1.despliegue - de - La - VidaDocumento3 páginasEnsayo 1.despliegue - de - La - VidaAlisson Camacho ErazoAún no hay calificaciones
- Desarrollo Sustentable o Ética AmbientalDocumento16 páginasDesarrollo Sustentable o Ética AmbientalLuis MaciasAún no hay calificaciones
- Yépez R. y Barret D. (1994) La Interdisciplinaridad, Lo Biologico y Lo Social en La Formación de MédicosDocumento20 páginasYépez R. y Barret D. (1994) La Interdisciplinaridad, Lo Biologico y Lo Social en La Formación de MédicosSandra L. HerranAún no hay calificaciones
- Reflexiones Sobre La Explicación Evolutiva en Ciencias CognitivasDocumento16 páginasReflexiones Sobre La Explicación Evolutiva en Ciencias CognitivasCarlosStevenAún no hay calificaciones
- Programa EvolucionDocumento6 páginasPrograma EvolucionTomas FernandezAún no hay calificaciones
- RESUMEN Yuni UrbanoDocumento6 páginasRESUMEN Yuni UrbanoGabriela di benedetto100% (3)
- Complejidad AF Tomo IDocumento337 páginasComplejidad AF Tomo IHéctor Reyes100% (4)
- La evolución del conocimiento científico: cómo construye su conocimiento la cienciaDe EverandLa evolución del conocimiento científico: cómo construye su conocimiento la cienciaAún no hay calificaciones
- El fracaso del desarrollo y la opción por el buen vivirDe EverandEl fracaso del desarrollo y la opción por el buen vivirAún no hay calificaciones
- Patrimonio ambiental y conocimiento local: Geografía de los actores socialesDe EverandPatrimonio ambiental y conocimiento local: Geografía de los actores socialesAún no hay calificaciones
- Un viaje por las alternativas al desarrollo: Perspectivas y propuestas teóricasDe EverandUn viaje por las alternativas al desarrollo: Perspectivas y propuestas teóricasAún no hay calificaciones
- Modulo 1Documento4 páginasModulo 1Elsie EstherAún no hay calificaciones
- Tarea Eca 1 Kristbel CaamañoDocumento1 páginaTarea Eca 1 Kristbel CaamañoElsie EstherAún no hay calificaciones
- Proyecto 3 Semana 1Documento1 páginaProyecto 3 Semana 1Elsie EstherAún no hay calificaciones
- Formato de Planificación de Proyectos Escolares-1Documento1 páginaFormato de Planificación de Proyectos Escolares-1Elsie EstherAún no hay calificaciones
- Inventos Del Siglo XVII y Siglo XVIIIDocumento6 páginasInventos Del Siglo XVII y Siglo XVIIIElsie EstherAún no hay calificaciones
- Proyecto Segundo SemestreDocumento157 páginasProyecto Segundo SemestreElsie EstherAún no hay calificaciones
- Hueso Temporal DiapositivasDocumento17 páginasHueso Temporal DiapositivasElsie Esther100% (1)
- Proyecto de Noviembre 1Documento3 páginasProyecto de Noviembre 1Elsie EstherAún no hay calificaciones
- Proyecto Segundo SemestreDocumento157 páginasProyecto Segundo SemestreElsie EstherAún no hay calificaciones
- Bioseguridad en Los Laboratorios ClinicosDocumento4 páginasBioseguridad en Los Laboratorios ClinicosElsie EstherAún no hay calificaciones
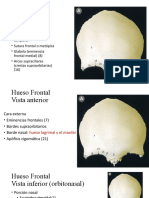
- Hueso Frontal DiapositivasDocumento6 páginasHueso Frontal DiapositivasElsie Esther100% (2)
- Hueso Occipital DiapositivasDocumento9 páginasHueso Occipital DiapositivasElsie EstherAún no hay calificaciones
- Tecnica InfraorbitariaDocumento1 páginaTecnica InfraorbitariaNatalia EstradaAún no hay calificaciones
- Artistas de La Región UcayaliDocumento16 páginasArtistas de La Región UcayaliJorge Rodríguez50% (2)
- Unidad 2 4to DSDocumento6 páginasUnidad 2 4to DSRENATO FABIAN ROBLES MATOSAún no hay calificaciones
- Lectura CríticaDocumento11 páginasLectura CríticaCristian Camilo Pava Martinez0% (2)
- Taller RequisitosDocumento4 páginasTaller RequisitosBoris Trujillo E.Aún no hay calificaciones
- Contaminación RadioactivaDocumento12 páginasContaminación RadioactivaAlvaro Diaz DelgadoAún no hay calificaciones
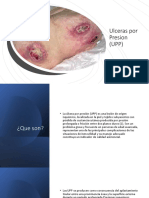
- Ulceras Por Presion (UPP)Documento10 páginasUlceras Por Presion (UPP)papugerapro2006Aún no hay calificaciones
- Casos ClinicosDocumento6 páginasCasos ClinicosMERLY MASILELI MENDEZ EULOGIOAún no hay calificaciones
- InfografíaDocumento1 páginaInfografíaBrayan RodriguezAún no hay calificaciones
- Hernia Discal - PlexoSolar26Documento13 páginasHernia Discal - PlexoSolar26ESMERDALINAAún no hay calificaciones
- MaferDocumento2 páginasMaferMafer AguilarAún no hay calificaciones
- Ta1 - Grupo 6Documento14 páginasTa1 - Grupo 6wcastro.lazaroAún no hay calificaciones
- Jugar Sopas de Letras de DESARROLLO HUMANO 41 en LíneaDocumento1 páginaJugar Sopas de Letras de DESARROLLO HUMANO 41 en Líneaelmer arrietaAún no hay calificaciones
- El Coronel No Tiene Quien Le EscribaDocumento2 páginasEl Coronel No Tiene Quien Le EscribaOsmar Alconz VadilloAún no hay calificaciones
- 7 Votos Del ExitoDocumento3 páginas7 Votos Del ExitoMarvel José Ardon FigueroaAún no hay calificaciones
- Telesur ¿Poder Blando Contrahegemónico? PDFDocumento175 páginasTelesur ¿Poder Blando Contrahegemónico? PDFhugosuppo@mac.comAún no hay calificaciones
- Permiso de Partenidad en FinlandiaDocumento3 páginasPermiso de Partenidad en FinlandiaLoreto CerdáAún no hay calificaciones
- Monografia TerminadaDocumento22 páginasMonografia TerminadaMirella Delgado EnriquezAún no hay calificaciones
- Art. 183.a. - Pornografia InfantilDocumento4 páginasArt. 183.a. - Pornografia InfantilMarco Aurelio Piña CarrascalAún no hay calificaciones
- Ejercicios DRXDocumento30 páginasEjercicios DRXNelson VelaAún no hay calificaciones
- GESTALTDocumento20 páginasGESTALTNicoll VicenteAún no hay calificaciones
- Lawrence Vale 12Documento32 páginasLawrence Vale 12Siglo21MediaGroupAún no hay calificaciones
- Ejercicios de Costos Por AbsorcionDocumento15 páginasEjercicios de Costos Por AbsorcionAbigail AlvasAún no hay calificaciones
- Resumen - RAICES PROFUNDASDocumento8 páginasResumen - RAICES PROFUNDASBelen FerreiraAún no hay calificaciones
- JUAN GOYTISOLO La Herencia de Cervantes 080196Documento3 páginasJUAN GOYTISOLO La Herencia de Cervantes 080196maxaub82Aún no hay calificaciones
- KANBANDocumento3 páginasKANBANjessica chungataAún no hay calificaciones
- Guía de Ortografía 1Documento5 páginasGuía de Ortografía 1Norelys BriceñoAún no hay calificaciones
- ENSAYO 5 El Docente y La Comunicación Didáctica en El Proceso de Enseñanza-AprendizajeDocumento4 páginasENSAYO 5 El Docente y La Comunicación Didáctica en El Proceso de Enseñanza-Aprendizajecardonarivera8780% (5)
- Los Organizadores GráficosDocumento9 páginasLos Organizadores Gráficosjlema2130Aún no hay calificaciones
- 3151-Texto Del Artículo-12279-1-10-20220128Documento28 páginas3151-Texto Del Artículo-12279-1-10-20220128economia urp2022Aún no hay calificaciones