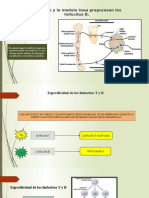
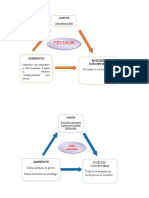
También podría gustarte
- 1 Ensayo Seleccion ClonalDocumento4 páginas1 Ensayo Seleccion ClonalPaola Luna0% (1)
- Biologia 5Documento94 páginasBiologia 5Santos Canahuire CondoriAún no hay calificaciones
- Funciones OPSONINASDocumento7 páginasFunciones OPSONINASCaro TorresAún no hay calificaciones
- Adquirida IIDocumento5 páginasAdquirida IIAlejandra UrrutiaAún no hay calificaciones
- Resumen 2 (Introducción A La Inmunología Humana)Documento17 páginasResumen 2 (Introducción A La Inmunología Humana)Jacqueline Agustina ToledoAún no hay calificaciones
- InmunologiaDocumento7 páginasInmunologiaAndres Felipe Prieto AlarconAún no hay calificaciones
- Copia de Inmunidad y AlergiaDocumento12 páginasCopia de Inmunidad y AlergiaDamian WayneAún no hay calificaciones
- Sistema Del ComplementoDocumento14 páginasSistema Del ComplementoDeysi LévanoAún no hay calificaciones
- Resumen Capitulo 2 4 Inmunologia Fundamentos Roitt 502947 Downloable 860800Documento20 páginasResumen Capitulo 2 4 Inmunologia Fundamentos Roitt 502947 Downloable 860800Michelle AmayaAún no hay calificaciones
- 5.clase 7 - 9 Inmunidad Innata IIIDocumento10 páginas5.clase 7 - 9 Inmunidad Innata IIIElías Damián Martínez TerénAún no hay calificaciones
- Chopita - Inmunidad InnataDocumento9 páginasChopita - Inmunidad InnataMartita JuárezAún no hay calificaciones
- Principios Basicos de InmunologiaDocumento58 páginasPrincipios Basicos de Inmunologiadrjaimerojas100% (7)
- Imprimir Agresion (1) - PagenumberDocumento135 páginasImprimir Agresion (1) - PagenumberSilvi OrregoAún no hay calificaciones
- Sistema Del ComplementoDocumento35 páginasSistema Del ComplementoRomina ArgumedoAún no hay calificaciones
- Bio PascuaDocumento3 páginasBio PascuaLucia MonsonisAún no hay calificaciones
- Inmunidad y AntibioticosDocumento13 páginasInmunidad y AntibioticosKrystel Volgger GarciaAún no hay calificaciones
- Inmunología Resumen ClasesDocumento19 páginasInmunología Resumen ClasesMajo MachadoAún no hay calificaciones
- Sistema de ComplementoDocumento1 páginaSistema de ComplementoAna Cristina Herrera ContrerasAún no hay calificaciones
- REACCIONES INMUNOLÓGICAS Sistema de Complemento RESUMENDocumento11 páginasREACCIONES INMUNOLÓGICAS Sistema de Complemento RESUMENIngrid Tatiana LozanoAún no hay calificaciones
- CitocinasDocumento3 páginasCitocinasalan fernando100% (1)
- PRACTICAS DE InmunologiaDocumento8 páginasPRACTICAS DE InmunologiaZuleima MesaAún no hay calificaciones
- Guía AyD Teoria 2do PrcialDocumento11 páginasGuía AyD Teoria 2do PrcialNicole FigueroaAún no hay calificaciones
- Cuestionario de Inmunologia CorreccionDocumento3 páginasCuestionario de Inmunologia CorreccionRonald D. CastroAún no hay calificaciones
- 0904 - Laboratorio Clinico 3Documento30 páginas0904 - Laboratorio Clinico 3Mayra RetamozoAún no hay calificaciones
- Bloque 1 Preguntas - Adrian Muñoz CastroDocumento7 páginasBloque 1 Preguntas - Adrian Muñoz CastroAdrian MuñozAún no hay calificaciones
- Examen ParcialDocumento4 páginasExamen ParcialFabi CunAún no hay calificaciones
- Resistencia Del Organismo A La InfeccionDocumento12 páginasResistencia Del Organismo A La InfeccionKarla AcuñaAún no hay calificaciones
- 5 Sistema de ComplementoDocumento40 páginas5 Sistema de ComplementoLuan NettoAún no hay calificaciones
- Sistema de Complemento, Vias y FuncionesDocumento6 páginasSistema de Complemento, Vias y FuncionesAdrian Briones100% (1)
- Ensayo ParasitologiaDocumento9 páginasEnsayo ParasitologiaANGEL VLADIMIR CHOLICO TORRESAún no hay calificaciones
- 1er Parcial ADNDocumento4 páginas1er Parcial ADNMaribel del CuraAún no hay calificaciones
- Segunda ParteDocumento8 páginasSegunda ParteSeccia Salgado ToledoAún no hay calificaciones
- Introduccion A La InmunologiaDocumento7 páginasIntroduccion A La InmunologiaPaula VelezAún no hay calificaciones
- Informe 5 Histología SeminarioDocumento11 páginasInforme 5 Histología SeminarioSadae BDAún no hay calificaciones
- Repaso Tema 4 InmunologiaDocumento8 páginasRepaso Tema 4 InmunologiaorlynmagdalysAún no hay calificaciones
- Sistema Inmune Innato y Adquirido. Respuesta Contra Bacterias, Virus y Parásitos. Clase 4Documento7 páginasSistema Inmune Innato y Adquirido. Respuesta Contra Bacterias, Virus y Parásitos. Clase 4marianaAún no hay calificaciones
- El Sistema Del ComplementoDocumento5 páginasEl Sistema Del ComplementoPAOLA ARCILAAún no hay calificaciones
- Clase 9-Inmunidad InnataDocumento11 páginasClase 9-Inmunidad Innataandreasposaro73Aún no hay calificaciones
- Inmunología Tema 2: La Respuesta Inmunitaria CelularDocumento10 páginasInmunología Tema 2: La Respuesta Inmunitaria CelularanaAún no hay calificaciones
- SEGUNDO CORTE Dana Complejo Mayor de HistocompatibilidadDocumento7 páginasSEGUNDO CORTE Dana Complejo Mayor de HistocompatibilidadwendyAún no hay calificaciones
- 6-Sistema de ComplementoDocumento9 páginas6-Sistema de Complementogeobannys cedeñoAún no hay calificaciones
- Anticuerpos MonoclonalesDocumento23 páginasAnticuerpos MonoclonalesFernando AkatsukiAún no hay calificaciones
- Los Linfocitos NK y Sus ReceptoresDocumento35 páginasLos Linfocitos NK y Sus Receptoreseliana callpaAún no hay calificaciones
- Cuestionario Del Sistema Del ComplementoDocumento6 páginasCuestionario Del Sistema Del ComplementoKarla BastidaAún no hay calificaciones
- Como Funciona El ComplementoDocumento2 páginasComo Funciona El ComplementoValeria Michelle Reyes GomezAún no hay calificaciones
- Resumen Inmuno 1eqDocumento3 páginasResumen Inmuno 1eqAndrés HerreraAún no hay calificaciones
- MediadoresDocumento15 páginasMediadoresAndres Le FontiAún no hay calificaciones
- Examen de InmunologiaDocumento10 páginasExamen de InmunologiaDante JosueAún no hay calificaciones
- InformesDocumento17 páginasInformesAndré MauricioAún no hay calificaciones
- Antigenos IntracelularesDocumento5 páginasAntigenos IntracelularesJennifer Stefania Meza MolinaAún no hay calificaciones
- Segunda Parte (Autoguardado)Documento14 páginasSegunda Parte (Autoguardado)Seccia Salgado ToledoAún no hay calificaciones
- Respuesta Celular y HumoralDocumento33 páginasRespuesta Celular y HumoralHugoRafaelGarciaMirandaAún no hay calificaciones
- Monitoria AnticuerposDocumento6 páginasMonitoria Anticuerpos20211211015Aún no hay calificaciones
- Inmunidad HumoralDocumento3 páginasInmunidad HumoralRobert Carrillo ZeñaAún no hay calificaciones
- Inmunidad HumoralDocumento7 páginasInmunidad HumoralMilagros FloresAún no hay calificaciones
- Generalidades Del Sistema InmunoDocumento7 páginasGeneralidades Del Sistema InmunoNICOLE MEJIA CEPEDAAún no hay calificaciones
- Monografía Bioquímica PDocumento14 páginasMonografía Bioquímica PMayfe CarreraAún no hay calificaciones
- Examen BiologiaDocumento7 páginasExamen BiologiaPedro García AguilarAún no hay calificaciones
- Sistema Inmune AdquiridoDocumento23 páginasSistema Inmune AdquiridoIrving100% (1)
- Trabajo de BiologíaDocumento4 páginasTrabajo de BiologíaKarla Pérez Zúñiga20% (5)
- Antisépticos Mas Utilizados en La Práctica Médica y Su Aplicación Práctica P1Documento3 páginasAntisépticos Mas Utilizados en La Práctica Médica y Su Aplicación Práctica P1JAMMILETH LENNY SALVATIERRA GUTIERREZAún no hay calificaciones
- Cuadro Sináptico Comparativo Sobre Celulitis y ErisipelaDocumento2 páginasCuadro Sináptico Comparativo Sobre Celulitis y ErisipelaJAMMILETH LENNY SALVATIERRA GUTIERREZAún no hay calificaciones
- Como Se Origina La Bilirrubina Indirecta y DirectaDocumento2 páginasComo Se Origina La Bilirrubina Indirecta y DirectaJAMMILETH LENNY SALVATIERRA GUTIERREZAún no hay calificaciones
- Tuberculosis PulmonarDocumento10 páginasTuberculosis PulmonarJAMMILETH LENNY SALVATIERRA GUTIERREZAún no hay calificaciones
- Síndrome Amotivacional Por CannabisDocumento3 páginasSíndrome Amotivacional Por CannabisJAMMILETH LENNY SALVATIERRA GUTIERREZAún no hay calificaciones
- Caso Clínico de Colecistitis Aguda P1Documento11 páginasCaso Clínico de Colecistitis Aguda P1JAMMILETH LENNY SALVATIERRA GUTIERREZAún no hay calificaciones
- BRONQUIECTASIADocumento8 páginasBRONQUIECTASIAJAMMILETH LENNY SALVATIERRA GUTIERREZAún no hay calificaciones
- Respuesta Inmune Ante Bacterias.Documento7 páginasRespuesta Inmune Ante Bacterias.JAMMILETH LENNY SALVATIERRA GUTIERREZAún no hay calificaciones
- Respuesta Inmunitaria Ante HongosDocumento3 páginasRespuesta Inmunitaria Ante HongosJAMMILETH LENNY SALVATIERRA GUTIERREZAún no hay calificaciones
- Respuesta Inmunitaria Ante ParásitosDocumento5 páginasRespuesta Inmunitaria Ante ParásitosJAMMILETH LENNY SALVATIERRA GUTIERREZAún no hay calificaciones
- Caso Clinico Sindrome Piramidal y ExtrapiramidalDocumento4 páginasCaso Clinico Sindrome Piramidal y ExtrapiramidalJAMMILETH LENNY SALVATIERRA GUTIERREZAún no hay calificaciones
- Reumatologia Historia Clinica de Artritis Septica y Artrosis p1Documento8 páginasReumatologia Historia Clinica de Artritis Septica y Artrosis p1JAMMILETH LENNY SALVATIERRA GUTIERREZAún no hay calificaciones
- Reumatología P1Documento3 páginasReumatología P1JAMMILETH LENNY SALVATIERRA GUTIERREZAún no hay calificaciones
- Células y Mecanismo de La Inmunidad InnataDocumento2 páginasCélulas y Mecanismo de La Inmunidad InnataJAMMILETH LENNY SALVATIERRA GUTIERREZAún no hay calificaciones
- Microorganismos Sistema NerviosoDocumento20 páginasMicroorganismos Sistema NerviosoPeter ValeriusAún no hay calificaciones
- Trabajo de Constitucional (RS) grupal-PROYECTO SOCIO EDUCATIVO "SENSIBILIZACIÒN SOBRE MEDIDAS PARA PREVENIR EL COVID-19 EN LA LIMPIEZA DEL HOGAR"Documento14 páginasTrabajo de Constitucional (RS) grupal-PROYECTO SOCIO EDUCATIVO "SENSIBILIZACIÒN SOBRE MEDIDAS PARA PREVENIR EL COVID-19 EN LA LIMPIEZA DEL HOGAR"Carmen Cánova Valladares67% (3)
- Idoc Consentimiento Prueba VihDocumento2 páginasIdoc Consentimiento Prueba VihPer HuberAún no hay calificaciones
- Seminario 3 - VIH, COVID-19 y VariantesDocumento42 páginasSeminario 3 - VIH, COVID-19 y VariantesFrings VarillasAún no hay calificaciones
- Protocolo Bioseguridad Covic (1) BarberiaDocumento24 páginasProtocolo Bioseguridad Covic (1) Barberiajuan esteban blandonAún no hay calificaciones
- Ficha Tecnica Test Covid-19Documento3 páginasFicha Tecnica Test Covid-19Emerson CorderoAún no hay calificaciones
- 201.131.125.54 19 Vitalabweb IndexDocumento1 página201.131.125.54 19 Vitalabweb IndexBruce CarmonaAún no hay calificaciones
- LIBROZOONOSIS2020UVISBN9786078490837Documento171 páginasLIBROZOONOSIS2020UVISBN9786078490837DANIEL VELA DIAZAún no hay calificaciones
- Catalogo de VirusDocumento32 páginasCatalogo de VirusJorge Agusto Luis VilcamizaAún no hay calificaciones
- Ensayo de La GripeDocumento5 páginasEnsayo de La GripeDiego EspinozaAún no hay calificaciones
- Seminario - Infecciones ViralesDocumento47 páginasSeminario - Infecciones ViralesLAURA NATALY PUYO FOREROAún no hay calificaciones
- Supuesto Practico 4 Nutricion EtapasDocumento5 páginasSupuesto Practico 4 Nutricion EtapasLF MonteroAún no hay calificaciones
- Ficha Tecnica Clindamicina PDFDocumento4 páginasFicha Tecnica Clindamicina PDFIVAN gomezAún no hay calificaciones
- Diagnóstico Serológico PDFDocumento38 páginasDiagnóstico Serológico PDFAndres Felipe Leal RodriguezAún no hay calificaciones
- Virus Del Mosaico de La Alfalfa YaDocumento16 páginasVirus Del Mosaico de La Alfalfa Yajulian rojas s.Aún no hay calificaciones
- Prevención de Riesgos Del Personal de SaludDocumento6 páginasPrevención de Riesgos Del Personal de Saludlise1013Aún no hay calificaciones
- Ficha Técnica DES-AMONDocumento5 páginasFicha Técnica DES-AMONAngela CavourAún no hay calificaciones
- Marco Teorico de MicroDocumento3 páginasMarco Teorico de MicroMimi MartinezAún no hay calificaciones
- Lienzo de La Propuesta de Valor - Semana 12Documento3 páginasLienzo de La Propuesta de Valor - Semana 12Cintia Leonela Huaman CuyubambaAún no hay calificaciones
- Biología 1Documento3 páginasBiología 1KARELYS ANHALYA INTRIAGO QUINDEAún no hay calificaciones
- Modelo de La Diseminación de Genes de Resistencia - En.esDocumento11 páginasModelo de La Diseminación de Genes de Resistencia - En.esPretty LitiñaAún no hay calificaciones
- CLASE 1-MICROB GeneralidadesDocumento59 páginasCLASE 1-MICROB Generalidadescarolina vegaAún no hay calificaciones
- Triada EpidemiologicaDocumento8 páginasTriada Epidemiologicanaysha50% (6)
- Multiclean Q-256 EspDocumento7 páginasMulticlean Q-256 EspJohncito ValAún no hay calificaciones
- Microbiologia de Los Alimentos Tarea 1Documento6 páginasMicrobiologia de Los Alimentos Tarea 1daniel gomezAún no hay calificaciones
- GUÍA#4-9 (27-30 de Abril)Documento6 páginasGUÍA#4-9 (27-30 de Abril)Departamento de Humanidades NCPAún no hay calificaciones
- 01.00.00 Cerco Perimetrico Tipo Uni - Mod - Plan CovidDocumento92 páginas01.00.00 Cerco Perimetrico Tipo Uni - Mod - Plan CovidCarlosAún no hay calificaciones
- MorfologiaDocumento12 páginasMorfologiabryan Martín ramirez MartínezAún no hay calificaciones
- Protocolo Covid-19 TipoDocumento11 páginasProtocolo Covid-19 TipoSneijder Animas100% (1)