También podría gustarte
- El cannabis en patologías del sistema nervioso centralDe EverandEl cannabis en patologías del sistema nervioso centralAún no hay calificaciones
- Fibras NerviosasDocumento4 páginasFibras NerviosasGUTIERREZ GARCIA ANET YARELIAún no hay calificaciones
- Somato SensorialDocumento6 páginasSomato SensorialcandyAún no hay calificaciones
- Neuro PRODocumento80 páginasNeuro PRONoe AlexanderAún no hay calificaciones
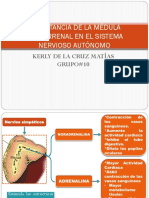
- Importancia de La Médula SuprarrenalDocumento10 páginasImportancia de La Médula SuprarrenalGlenda AracelyAún no hay calificaciones
- Neurofisiología VÍAS SENSITIVAS-2Documento34 páginasNeurofisiología VÍAS SENSITIVAS-2claudiamartinez1011Aún no hay calificaciones
- Información SensorialDocumento99 páginasInformación Sensorialnelly fernandaAún no hay calificaciones
- Fisiologiasensorial 091021195921 Phpapp01Documento38 páginasFisiologiasensorial 091021195921 Phpapp01Josue RojadAún no hay calificaciones
- Sensibilidad 1Documento26 páginasSensibilidad 1Luna ValenciaAún no hay calificaciones
- 2565 BT 08 19 ReceptoresDocumento24 páginas2565 BT 08 19 ReceptoresRodrigoAún no hay calificaciones
- Tabla Células FISIO - AMM - DEMR - DRCDocumento5 páginasTabla Células FISIO - AMM - DEMR - DRCREYNALDO JESUS HERNANDEZ RODRIGUEZAún no hay calificaciones
- Vias Sensitivas GeneralesDocumento32 páginasVias Sensitivas Generalesmahlyceu2022Aún no hay calificaciones
- Conferencia Semana 7 PDFDocumento99 páginasConferencia Semana 7 PDFĐäřêł ĞŤøwėřşAún no hay calificaciones
- Integración Sensorial CompletoDocumento8 páginasIntegración Sensorial CompletoYuliana Victoria BARRIONUEVO CARPIOAún no hay calificaciones
- Fisiologia - CLASE - Somatosensorial 2017Documento51 páginasFisiologia - CLASE - Somatosensorial 2017Stania CamposAún no hay calificaciones
- Semana 1 Sistema SensorialDocumento12 páginasSemana 1 Sistema SensorialLuis HerediaAún no hay calificaciones
- Unidad 3 Sistema Sensitivo y Motor. Equipo 5 Grupo 45686Documento210 páginasUnidad 3 Sistema Sensitivo y Motor. Equipo 5 Grupo 45686BhanuDeviDasiAún no hay calificaciones
- CAP46Documento2 páginasCAP46Juan Jose RUIZAún no hay calificaciones
- Clase N°4 Enfermeria - Somatosensorial y Dolor 2021Documento37 páginasClase N°4 Enfermeria - Somatosensorial y Dolor 2021Estefania conejerosAún no hay calificaciones
- Sesión 6 202110 Sistema Somatosensorial.Documento24 páginasSesión 6 202110 Sistema Somatosensorial.Antonia Samur ContrerasAún no hay calificaciones
- Fisiologia Nerviosa. Guía.Documento42 páginasFisiologia Nerviosa. Guía.Marines BarretoAún no hay calificaciones
- Los Estímulos y Receptores SensorialesDocumento1 páginaLos Estímulos y Receptores SensorialesDixi PixiAún no hay calificaciones
- Fisiología SN 03Documento44 páginasFisiología SN 03Vipex10Aún no hay calificaciones
- Funciones Sensoriales Del Sistema Nervioso 2018-IIDocumento39 páginasFunciones Sensoriales Del Sistema Nervioso 2018-IIAlvaro LozanoAún no hay calificaciones
- Fisiologia SensorialDocumento85 páginasFisiologia SensorialEricksson Moran100% (2)
- Receptores Sensoriales 2019Documento92 páginasReceptores Sensoriales 2019Yulisa RojasAún no hay calificaciones
- 04b Neurofisiologia SensorialDocumento99 páginas04b Neurofisiologia SensorialAbel D. PacoAún no hay calificaciones
- Receptores Sensoriales .PotDocumento3 páginasReceptores Sensoriales .Potraquel najera0% (1)
- 06 Sistemas SensorialesDocumento60 páginas06 Sistemas SensorialesMister ItoldyaAún no hay calificaciones
- Clase Sensibilidad Somatica-178 PDFDocumento46 páginasClase Sensibilidad Somatica-178 PDFMaría Catalina BeneventiAún no hay calificaciones
- Sistema Vegatito, Sensitivo, MotorDocumento103 páginasSistema Vegatito, Sensitivo, MotorXimena MarilúAún no hay calificaciones
- Semana 1 VacacionalDocumento40 páginasSemana 1 VacacionalLuis HerediaAún no hay calificaciones
- Repaso Parcial Fisio 220807 105144Documento319 páginasRepaso Parcial Fisio 220807 105144Adriana NeyraAún no hay calificaciones
- Receptores y Efectores PDFDocumento24 páginasReceptores y Efectores PDFcamiAún no hay calificaciones
- Sensibilidad UpaoDocumento50 páginasSensibilidad UpaoYhojana UrbanoAún no hay calificaciones
- Fisiologia SensorialDocumento99 páginasFisiologia SensorialLuis Herrera100% (1)
- Cap 47 Ecpetores Sensitivos y Circuitos NeuronalesDocumento9 páginasCap 47 Ecpetores Sensitivos y Circuitos NeuronalesdavidmoralesAún no hay calificaciones
- Tema 29 (Terminales Nerviosas)Documento14 páginasTema 29 (Terminales Nerviosas)Carlos Daniel Cárdenas SantosAún no hay calificaciones
- ReceptoresDocumento40 páginasReceptoresFelipe Martínez Arbeláez100% (1)
- Capítulo 47 Fisiología Guyton y Hall 14 Ed.Documento9 páginasCapítulo 47 Fisiología Guyton y Hall 14 Ed.Manuela MenendezAún no hay calificaciones
- Receptores SensorialesDocumento29 páginasReceptores SensorialesSamuel Salar PadillaAún no hay calificaciones
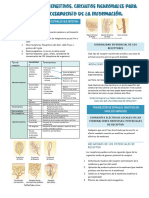
- Receptores Sensitivos, Circuitos Neuronales para El Procesamiento de La Información y Sensibilidad SomáticaDocumento3 páginasReceptores Sensitivos, Circuitos Neuronales para El Procesamiento de La Información y Sensibilidad SomáticaMayra G Sanchez LeonardoAún no hay calificaciones
- CAPÍTULO 47 - Fisiología GUYTONDocumento3 páginasCAPÍTULO 47 - Fisiología GUYTONRashel MoscosoAún no hay calificaciones
- La Sensibilidad y El DolorDocumento31 páginasLa Sensibilidad y El DolorAlex QuinteroAún no hay calificaciones
- Receptores SensorialesDocumento12 páginasReceptores SensorialesSharon_n-AnnyeongAún no hay calificaciones
- Funciones Sensoriales Del SNDocumento34 páginasFunciones Sensoriales Del SNCe SecoAún no hay calificaciones
- Capitulo 4 Procesos SensorialesDocumento48 páginasCapitulo 4 Procesos SensorialesFERNANDO RAMOSAún no hay calificaciones
- Fisiologia SensorialDocumento95 páginasFisiologia SensorialJunior RamírezAún no hay calificaciones
- Biofísica Sensorial Ondas Sonoras y ElectromagnéticasDocumento42 páginasBiofísica Sensorial Ondas Sonoras y ElectromagnéticasEsmeralda CruzAún no hay calificaciones
- Resumen 3 - Receptores de La Información y Conducción Del Pulso NerviosoDocumento14 páginasResumen 3 - Receptores de La Información y Conducción Del Pulso NerviosoSaid Izurieta G.Aún no hay calificaciones
- ByNF - Teórico ER - GalottoDocumento45 páginasByNF - Teórico ER - GalottoAnaliaSosaAún no hay calificaciones
- Fisiologia Percepcion SensorialDocumento61 páginasFisiologia Percepcion SensorialAndrea Cadillo PomaAún no hay calificaciones
- PRACTICA NO. 6-Receptores-Sensoriales EQ 5Documento12 páginasPRACTICA NO. 6-Receptores-Sensoriales EQ 5Joguard Lofcraft100% (1)
- CAPÍTULO 8 - Neurotransmisión Somatosensitiva - Tacto, Dolor y TemperaturaDocumento25 páginasCAPÍTULO 8 - Neurotransmisión Somatosensitiva - Tacto, Dolor y TemperaturaMJ ArgentoAún no hay calificaciones
- Receptores Sensitivos 2Documento12 páginasReceptores Sensitivos 2kamiila TrujilloAún no hay calificaciones
- FISIOLOGÍA SENSORIAL DiaposDocumento39 páginasFISIOLOGÍA SENSORIAL DiaposlizethgomezcarlinAún no hay calificaciones
- Ficha Resumen Del Documento RIVASDocumento10 páginasFicha Resumen Del Documento RIVASMau XoloAún no hay calificaciones
- MORFOFISIOLOGÍA I - Fisiología Semana 1: Dra. Lissett Jeanette Fernández RodríguezDocumento74 páginasMORFOFISIOLOGÍA I - Fisiología Semana 1: Dra. Lissett Jeanette Fernández RodríguezGESU PIERO LOPEZ MEJIAAún no hay calificaciones
- Sesión 6 Receptores y Vías Sensitivas 22092022Documento22 páginasSesión 6 Receptores y Vías Sensitivas 22092022jeihop02Aún no hay calificaciones
- SensibilidadDocumento50 páginasSensibilidademilcita2011Aún no hay calificaciones
- EmergenciasDocumento21 páginasEmergenciasPosting Yourself RDAún no hay calificaciones
- Introducción A Las CienciasDocumento3 páginasIntroducción A Las Cienciasraul lugoAún no hay calificaciones
- Diptico DanielaDocumento2 páginasDiptico DanielaJuvenal Alfredo Huerta ZoluagaAún no hay calificaciones
- Niveles de Organización Estructural. JuanDocumento2 páginasNiveles de Organización Estructural. JuanJUAN SEBASTIAN MEDINA BELTRÁNAún no hay calificaciones
- Folleto de Mes PDFDocumento36 páginasFolleto de Mes PDFDeyadhiraTovarAún no hay calificaciones
- Sex Beyond The Genitalia - The Human Brain Mosaic (1) .En - EsDocumento6 páginasSex Beyond The Genitalia - The Human Brain Mosaic (1) .En - EsLeFührer Zabdiel PazaranAún no hay calificaciones
- Actividad 16Documento4 páginasActividad 16david rAún no hay calificaciones
- Fase 2 Fisiologia VegetalDocumento7 páginasFase 2 Fisiologia VegetaljuliedAún no hay calificaciones
- Práctica Extra Diseccion CorazonDocumento4 páginasPráctica Extra Diseccion CorazonPAULO RODRIGUEZ RECHYAún no hay calificaciones
- Clases de Bases de Anatomia y FisiologiaDocumento271 páginasClases de Bases de Anatomia y FisiologiaJudith Stefany Condorcallo CamaAún no hay calificaciones
- Aparato de GolgiDocumento14 páginasAparato de GolgiFernando IxcotAún no hay calificaciones
- Introducción: T.12 Las Plantas SuperioresDocumento9 páginasIntroducción: T.12 Las Plantas SuperioresRDAún no hay calificaciones
- Mapa Conceptual GlandulasDocumento1 páginaMapa Conceptual GlandulasTania PeraltaAún no hay calificaciones
- 4.2. ENDOMEMBRANAS - El Complejo de GolgiDocumento31 páginas4.2. ENDOMEMBRANAS - El Complejo de GolgiMaria GarciaAún no hay calificaciones
- Tema 3Documento6 páginasTema 3Xisco Lozano SalvaAún no hay calificaciones
- Sistema NerviosoDocumento17 páginasSistema Nerviosoangeles lozano arevaloAún no hay calificaciones
- Sistema Digestivo en Animales PoligastricosDocumento8 páginasSistema Digestivo en Animales PoligastricosmarilynAún no hay calificaciones
- Líquidos Del OrganismoDocumento22 páginasLíquidos Del OrganismoHecdimar MalpicaAún no hay calificaciones
- Introduccion A La Biologia 2020 4to VirtualDocumento12 páginasIntroduccion A La Biologia 2020 4to VirtualYasmani LlanquiAún no hay calificaciones
- Informe Laboratorio Celulas Animales y VegetalesDocumento9 páginasInforme Laboratorio Celulas Animales y VegetalesDaniela Osorio QuirozAún no hay calificaciones
- Factores Que Influyen en El CrecimientoDocumento4 páginasFactores Que Influyen en El Crecimientoangel226293Aún no hay calificaciones
- Bioquímica de La Transmisión NerviosaDocumento4 páginasBioquímica de La Transmisión NerviosaKROSKIDOCAún no hay calificaciones
- Clase 6 Semana 4 - Reproducción en Cuyes - Aparato Reproductor Hembra y Macho - PubertadDocumento16 páginasClase 6 Semana 4 - Reproducción en Cuyes - Aparato Reproductor Hembra y Macho - PubertadDiandra Romero MoránAún no hay calificaciones
- Guia Sistema Nervioso. SantillanaDocumento12 páginasGuia Sistema Nervioso. SantillanaPedro Nelgui Vargas QuinteroAún no hay calificaciones
- Album IlustrativoDocumento15 páginasAlbum IlustrativoKevin GarciaAún no hay calificaciones
- Desarrollo de La PlacentaDocumento17 páginasDesarrollo de La Placentasamuelcolmenarestoledo2100% (1)
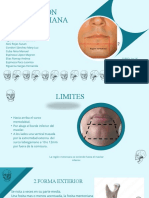
- Region MentonianaDocumento11 páginasRegion MentonianaSarai AcHáAún no hay calificaciones
- Electroestimulador V3Documento9 páginasElectroestimulador V3Julio CamposAún no hay calificaciones
- Tratamiento Conservador ESDocumento149 páginasTratamiento Conservador ESDiana Mallma BendezuAún no hay calificaciones
- Respuestas de Examen Olimpiada de Biología 2020Documento15 páginasRespuestas de Examen Olimpiada de Biología 2020Minchani BorreguinAún no hay calificaciones
- Recupera tu mente, reconquista tu vidaDe EverandRecupera tu mente, reconquista tu vidaCalificación: 5 de 5 estrellas5/5 (9)
- Psicología oscura: Una guía esencial de persuasión, manipulación, engaño, control mental, negociación, conducta humana, PNL y guerra psicológicaDe EverandPsicología oscura: Una guía esencial de persuasión, manipulación, engaño, control mental, negociación, conducta humana, PNL y guerra psicológicaCalificación: 4.5 de 5 estrellas4.5/5 (766)
- El poder del optimismo: Herramientas para vivir de forma más positivaDe EverandEl poder del optimismo: Herramientas para vivir de forma más positivaCalificación: 5 de 5 estrellas5/5 (16)
- Cómo hacer que te pasen cosas buenas: Entiende tu cerebro, gestiona tus emociones, mejora tu vidaDe EverandCómo hacer que te pasen cosas buenas: Entiende tu cerebro, gestiona tus emociones, mejora tu vidaCalificación: 5 de 5 estrellas5/5 (1877)
- La revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaDe EverandLa revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaCalificación: 5 de 5 estrellas5/5 (205)
- Tus Zonas Erroneas: Guía Para Combatir las Causas de la InfelicidadDe EverandTus Zonas Erroneas: Guía Para Combatir las Causas de la InfelicidadCalificación: 4.5 de 5 estrellas4.5/5 (1834)
- Yo Pude, ¡Tú Puedes!: Cómo tomar el control de tu bienestar emocional y convertirte en una persona imparable (edición revisada y expandida)De EverandYo Pude, ¡Tú Puedes!: Cómo tomar el control de tu bienestar emocional y convertirte en una persona imparable (edición revisada y expandida)Calificación: 5 de 5 estrellas5/5 (11)
- DMT: La molécula del espíritu (DMT: The Spirit Molecule): Las revolucionarias investigaciones de un medico sobre la biologia de las experiencias misticas y cercanas a la muerteDe EverandDMT: La molécula del espíritu (DMT: The Spirit Molecule): Las revolucionarias investigaciones de un medico sobre la biologia de las experiencias misticas y cercanas a la muerteCalificación: 4.5 de 5 estrellas4.5/5 (19)
- La madre emocionalmente ausente: Como reconocer y sanar los efectos invisibles del abandono emocional infantilDe EverandLa madre emocionalmente ausente: Como reconocer y sanar los efectos invisibles del abandono emocional infantilAún no hay calificaciones
- Las 6 necesidades de cada niño: Empoderar a padres e hijos a través de la ciencia de la conexiónDe EverandLas 6 necesidades de cada niño: Empoderar a padres e hijos a través de la ciencia de la conexiónCalificación: 4.5 de 5 estrellas4.5/5 (7)
- Cómo Conversar Con Cualquier Persona: Mejora tus habilidades sociales, desarrolla tu carisma, domina las conversaciones triviales y conviértete en una persona sociable para hacer verdaderos amigos y construir relaciones significativas.De EverandCómo Conversar Con Cualquier Persona: Mejora tus habilidades sociales, desarrolla tu carisma, domina las conversaciones triviales y conviértete en una persona sociable para hacer verdaderos amigos y construir relaciones significativas.Calificación: 5 de 5 estrellas5/5 (54)
- Resetea tu mente. Descubre de lo que eres capazDe EverandResetea tu mente. Descubre de lo que eres capazCalificación: 5 de 5 estrellas5/5 (196)
- La violencia invisible: Identificar, entender y superar la violencia psicológica que sufrimos (y ejercemos) en nuestra vida cotidianaDe EverandLa violencia invisible: Identificar, entender y superar la violencia psicológica que sufrimos (y ejercemos) en nuestra vida cotidianaCalificación: 4 de 5 estrellas4/5 (2)
- Cómo terminar lo que empiezas: El arte de perseverar, pasar a la acción, ejecutar los planes y tener disciplinaDe EverandCómo terminar lo que empiezas: El arte de perseverar, pasar a la acción, ejecutar los planes y tener disciplinaCalificación: 4.5 de 5 estrellas4.5/5 (6)
- Una mente en calma: Técnicas para manejar los pensamientos intrusivosDe EverandUna mente en calma: Técnicas para manejar los pensamientos intrusivosCalificación: 4.5 de 5 estrellas4.5/5 (145)
- El concepto Mulligan de terapia manual (Color)De EverandEl concepto Mulligan de terapia manual (Color)Calificación: 5 de 5 estrellas5/5 (3)
- Educar la atención: Cómo entrenar esta habilidad en niños y adultosDe EverandEducar la atención: Cómo entrenar esta habilidad en niños y adultosCalificación: 4.5 de 5 estrellas4.5/5 (10)
- El fin de la Ansiedad y los MiedosDe EverandEl fin de la Ansiedad y los MiedosCalificación: 4.5 de 5 estrellas4.5/5 (2)
- Ejercicios de Psicología Positiva para aumentar tu felicidadDe EverandEjercicios de Psicología Positiva para aumentar tu felicidadCalificación: 4.5 de 5 estrellas4.5/5 (18)
- Lee a las personas como un libro: Cómo analizar, entender y predecir las emociones, los pensamientos, las intenciones y los comportamientos de las personasDe EverandLee a las personas como un libro: Cómo analizar, entender y predecir las emociones, los pensamientos, las intenciones y los comportamientos de las personasCalificación: 4.5 de 5 estrellas4.5/5 (3)
- Homo antecessor: El nacimiento de una especieDe EverandHomo antecessor: El nacimiento de una especieCalificación: 5 de 5 estrellas5/5 (1)
- Psicología del Dinero - Comportamiento Humano y su Relación entre el Dinero y Felicidad.Domina tu Mente y las Emociones para Administrar mejor tus Finanzas.Mercados FinancierosDe EverandPsicología del Dinero - Comportamiento Humano y su Relación entre el Dinero y Felicidad.Domina tu Mente y las Emociones para Administrar mejor tus Finanzas.Mercados FinancierosCalificación: 5 de 5 estrellas5/5 (12)