También podría gustarte
- Determinación de M.O. en AguaDocumento8 páginasDeterminación de M.O. en AguaRonald AvilaAún no hay calificaciones
- Determinación Electroquímica de Plomo y Cadmio en Aguas SuperficialesDocumento8 páginasDeterminación Electroquímica de Plomo y Cadmio en Aguas SuperficialesDiego Alderete CuniberttiAún no hay calificaciones
- Determinacion de PH y Acidez TitulableDocumento25 páginasDeterminacion de PH y Acidez TitulableLuisa Fernanda Carvajal RamirezAún no hay calificaciones
- Oxigeno DisueltoDocumento4 páginasOxigeno DisueltobucanereAún no hay calificaciones
- TP0351 Calcio y Magnesio en Agua Con EDTA Por VolumetríaDocumento10 páginasTP0351 Calcio y Magnesio en Agua Con EDTA Por VolumetríasaphkielAún no hay calificaciones
- APLICACIONES de Laboratorio Practicas 5-9 Equilibrio y CineticaDocumento14 páginasAPLICACIONES de Laboratorio Practicas 5-9 Equilibrio y CineticaAxel SamaAún no hay calificaciones
- La Medida de PH y El Electrodo de VidrioDocumento4 páginasLa Medida de PH y El Electrodo de VidrioKelly PrioloAún no hay calificaciones
- Analisis de NitratosDocumento18 páginasAnalisis de NitratosEdgar BorjaAún no hay calificaciones
- Cuantificacion de Proteinas BestiaragDocumento29 páginasCuantificacion de Proteinas BestiaragGuillermo RTAún no hay calificaciones
- Sulfatos en Agua (Espectrofotométrico)Documento12 páginasSulfatos en Agua (Espectrofotométrico)tonomontiAún no hay calificaciones
- 2 Determinacion de CD PDFDocumento8 páginas2 Determinacion de CD PDFLuis Martin Escalante PillacaAún no hay calificaciones
- Practica de Velocidad de ReaccionDocumento12 páginasPractica de Velocidad de ReaccionJose Maria Quintas GironAún no hay calificaciones
- Determinación de Arsénico en Agua Por Espectrometría de Absorción AtómicaDocumento4 páginasDeterminación de Arsénico en Agua Por Espectrometría de Absorción AtómicaDany Daniel Oblitas SoteloAún no hay calificaciones
- Determinación de Potenciométrica de Cloruro en Orina Humana y Suero FisiológicoDocumento5 páginasDeterminación de Potenciométrica de Cloruro en Orina Humana y Suero FisiológicoJoan Stiven Astudillo VAún no hay calificaciones
- LABO DE REA Preinforme 2Documento7 páginasLABO DE REA Preinforme 2Adriana RosasAún no hay calificaciones
- Os Métodos Potenciométricos Se Basan en La Medición Del Potencial en Una Celda Electroquímica Sin Paso de Corriente ApreciableDocumento3 páginasOs Métodos Potenciométricos Se Basan en La Medición Del Potencial en Una Celda Electroquímica Sin Paso de Corriente ApreciablearnoldAún no hay calificaciones
- LABORATORIO #8 PrácticoDocumento6 páginasLABORATORIO #8 PrácticoNataly FernandaAún no hay calificaciones
- Practica 1 CineticaDocumento16 páginasPractica 1 CineticaLuiis AlbertooAún no hay calificaciones
- Magnesium CPZ SP WienerDocumento12 páginasMagnesium CPZ SP Wienerlupita riveraAún no hay calificaciones
- Métodos de Prueba Estándar para Calcio y Magnesio en Agua - Astm D511Documento8 páginasMétodos de Prueba Estándar para Calcio y Magnesio en Agua - Astm D511milidarksAún no hay calificaciones
- Informe6 QA DQODocumento4 páginasInforme6 QA DQOAdriana LlanoAún no hay calificaciones
- Informe de Laboratorio Redox PDFDocumento9 páginasInforme de Laboratorio Redox PDFStefany ÑungoAún no hay calificaciones
- Clase 2 Quimica AnaliticaDocumento55 páginasClase 2 Quimica AnaliticaWilder David Escalante Alarcon100% (1)
- Nom-117-Ssa1-1994 Plomo Arsenico Mercurio CadmioDocumento28 páginasNom-117-Ssa1-1994 Plomo Arsenico Mercurio CadmioricardojbellosocrjbcAún no hay calificaciones
- Articulo Cientifico. Procedimiento de Validación de Un Método para Cuantificar Mercurio en Sangre Por Espectroscopía de Absorción Atómica Con Descomposición Térmica y AmalgamaciónDocumento18 páginasArticulo Cientifico. Procedimiento de Validación de Un Método para Cuantificar Mercurio en Sangre Por Espectroscopía de Absorción Atómica Con Descomposición Térmica y AmalgamaciónTeFany Csc MousesitaAún no hay calificaciones
- Determinación de Cobre Por Potenciometria de Adicion EstandarDocumento8 páginasDeterminación de Cobre Por Potenciometria de Adicion EstandarAura Ballesteros MontealegreAún no hay calificaciones
- Informe #2-Oxidación Del Yoduro Con PersulfatoDocumento16 páginasInforme #2-Oxidación Del Yoduro Con PersulfatoJuniorRomeroAún no hay calificaciones
- Compuestos Nitrogenados No Proteicos - Grupo 2Documento9 páginasCompuestos Nitrogenados No Proteicos - Grupo 2Jessenia VásquezAún no hay calificaciones
- Agilent 7500 ICP-MS MauroDocumento3 páginasAgilent 7500 ICP-MS MauroYennadim Medina RealesAún no hay calificaciones
- Práctica de Titulación PotenciometricaDocumento11 páginasPráctica de Titulación PotenciometricaIrving Sakay Garcia MendoAún no hay calificaciones
- Marco TeoricoDocumento15 páginasMarco TeoricoYudi CameroAún no hay calificaciones
- SelenioDocumento2 páginasSelenioJonathan López0% (1)
- Informe Cruz, Meneses, NarvaezDocumento6 páginasInforme Cruz, Meneses, NarvaezSara Gabriela CGAún no hay calificaciones
- Practica Lab 8. Determinacion Espectrofotometrica UV - VIS Del FeDocumento10 páginasPractica Lab 8. Determinacion Espectrofotometrica UV - VIS Del FeKeyla QuezadaAún no hay calificaciones
- PRÁCTICA No1 (Azucares Reductores)Documento4 páginasPRÁCTICA No1 (Azucares Reductores)JuanDavidOrtegaOlaveAún no hay calificaciones
- Determinación Electroquímica de Plomo y Cadmio en AguasDocumento12 páginasDeterminación Electroquímica de Plomo y Cadmio en AguasDonna Granja RizzoAún no hay calificaciones
- PH en Aguas Por ElectrometriaDocumento7 páginasPH en Aguas Por ElectrometriaDaniel Orozco NaranjoAún no hay calificaciones
- Fer Color Aa Liquida SPDocumento3 páginasFer Color Aa Liquida SPcarina_gutiAún no hay calificaciones
- Validación Del Método de Determinación de Mercurio en Muestras de Sedimentos y Tejidos Biológicos Utilizando Un Analizador Directo de MercurioDocumento5 páginasValidación Del Método de Determinación de Mercurio en Muestras de Sedimentos y Tejidos Biológicos Utilizando Un Analizador Directo de MercurioDiego Alderete CuniberttiAún no hay calificaciones
- P4. Cinética de Halogenación de La AcetonaDocumento8 páginasP4. Cinética de Halogenación de La AcetonajairoSKATEMAN100% (1)
- Determinacion de Hierro Por Indicador y PotenciometricamenteDocumento9 páginasDeterminacion de Hierro Por Indicador y PotenciometricamenteLizeth Leon Chavez100% (1)
- Potenciometria y Sus Aplicaciones en El Campo de La CienciaDocumento5 páginasPotenciometria y Sus Aplicaciones en El Campo de La CienciaAyazo Lopez DaladielAún no hay calificaciones
- Informe de Laboratorio Redox PDFDocumento9 páginasInforme de Laboratorio Redox PDFOrlando Alvarez Cubillos0% (1)
- Optimizacion Tecnica de Arsenico Con Generador de HidrurosDocumento4 páginasOptimizacion Tecnica de Arsenico Con Generador de HidrurosAbi Ponce MenaAún no hay calificaciones
- Gastro RalDocumento2 páginasGastro RalRomario de la CruzAún no hay calificaciones
- 6330 Glicemia Enzimatica SP PDFDocumento3 páginas6330 Glicemia Enzimatica SP PDFJuan Flores OlguinAún no hay calificaciones
- Informe 3Documento9 páginasInforme 3Mishell Madelei Palacios HuatucoAún no hay calificaciones
- Informe Experimental Ag-CuDocumento8 páginasInforme Experimental Ag-CuEuclides GarciaAún no hay calificaciones
- Determinacion de Oxigeno Disuelto Por ElectroquimicaDocumento4 páginasDeterminacion de Oxigeno Disuelto Por ElectroquimicaBearKiarAún no hay calificaciones
- 4500-O-G. Método de La Membrana-Electrodo - ODDocumento6 páginas4500-O-G. Método de La Membrana-Electrodo - ODOperaciones L y LAún no hay calificaciones
- EnviarDocumento16 páginasEnviaresneider linares romeroAún no hay calificaciones
- DIEGODocumento23 páginasDIEGODiego Padilla SantosAún no hay calificaciones
- Universidad de Córdoba INFORME 7Documento8 páginasUniversidad de Córdoba INFORME 7Laura Carmona.Aún no hay calificaciones
- Biosensores GrafenoDocumento9 páginasBiosensores GrafenoDavid Zamora RubioAún no hay calificaciones
- Demanda Química de OxígenoDocumento11 páginasDemanda Química de OxígenoDafne AilynAún no hay calificaciones
- Determinacion de PH y Acidez TitulableDocumento25 páginasDeterminacion de PH y Acidez TitulableJorge Cortez ElizaldeAún no hay calificaciones
- Manual práctico de ensayos de toxicidad en medio acuático con organismos del género DaphniaDe EverandManual práctico de ensayos de toxicidad en medio acuático con organismos del género DaphniaCalificación: 1 de 5 estrellas1/5 (1)
- Espectrometría de masas. Interpretación de espectrosDe EverandEspectrometría de masas. Interpretación de espectrosAún no hay calificaciones
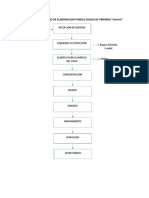
- A) OrganigramaDocumento1 páginaA) OrganigramaDarwin AlcivarAún no hay calificaciones
- Formas de Entrada en El M I - Canales de DistribuciònDocumento29 páginasFormas de Entrada en El M I - Canales de DistribuciònDarwin AlcivarAún no hay calificaciones
- Arcsa - LaboratorioDocumento3 páginasArcsa - LaboratorioDarwin AlcivarAún no hay calificaciones
- Espectroscopia Fotoelectronica de Rayos-X XPS en LDocumento8 páginasEspectroscopia Fotoelectronica de Rayos-X XPS en LDarwin AlcivarAún no hay calificaciones
- Senadi DG 2021 0197 ExDocumento1 páginaSenadi DG 2021 0197 ExDarwin Alcivar100% (1)
- Base de Oec Por Rte Inen 2021 05 27 ADocumento3 páginasBase de Oec Por Rte Inen 2021 05 27 ADarwin AlcivarAún no hay calificaciones
- Empacadoras Por PedalDocumento2 páginasEmpacadoras Por PedalDarwin AlcivarAún no hay calificaciones
- BPM InstalcionesDocumento25 páginasBPM InstalcionesDarwin AlcivarAún no hay calificaciones
- Empresas Certificadas BPM Plantas Procesadoras de Alimentos 30 Nov 2021Documento1628 páginasEmpresas Certificadas BPM Plantas Procesadoras de Alimentos 30 Nov 2021Darwin AlcivarAún no hay calificaciones
- Registro Unico de MYpimesDocumento1 páginaRegistro Unico de MYpimesDarwin AlcivarAún no hay calificaciones
- Empresas Certificadas BPM Plantas Procesadoras de Alimentos 31 Jul 2021Documento2291 páginasEmpresas Certificadas BPM Plantas Procesadoras de Alimentos 31 Jul 2021Darwin AlcivarAún no hay calificaciones
- PROFORMA2020STD72Documento1 páginaPROFORMA2020STD72Darwin AlcivarAún no hay calificaciones
- Proforma Dz400eDocumento2 páginasProforma Dz400eDarwin AlcivarAún no hay calificaciones
- TareaDocumento10 páginasTareaDarwin AlcivarAún no hay calificaciones
- Foro DebateDocumento5 páginasForo DebateDarwin AlcivarAún no hay calificaciones
- Habilidades DirectivasDocumento8 páginasHabilidades DirectivasDarwin AlcivarAún no hay calificaciones
- Ie B.3.1.2 Ali 03 - Sistematización Del Proceso de Buenas Prácticas de Alimentos Procesados - V2Documento77 páginasIe B.3.1.2 Ali 03 - Sistematización Del Proceso de Buenas Prácticas de Alimentos Procesados - V2Darwin AlcivarAún no hay calificaciones
- La Trazabilidad Contexto InternacionalDocumento27 páginasLa Trazabilidad Contexto InternacionalDarwin AlcivarAún no hay calificaciones
- Diagrama de FlujoDocumento3 páginasDiagrama de FlujoDarwin AlcivarAún no hay calificaciones