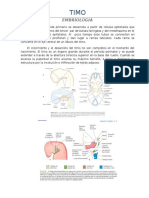
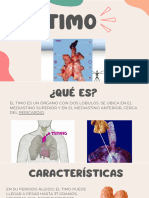
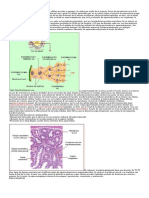
También podría gustarte
- Manual de Casos Prácticos de Derecho Civil - Nodrm 1Documento296 páginasManual de Casos Prácticos de Derecho Civil - Nodrm 1Macarena Ortali93% (40)
- Manual de Derecho Penal Chileno. Parte General - 2 Edición - NodrmDocumento805 páginasManual de Derecho Penal Chileno. Parte General - 2 Edición - NodrmM79% (24)
- Causales Del Despido Disciplinario en ChileDocumento385 páginasCausales Del Despido Disciplinario en ChileM100% (5)
- Biorreactores Unidad 4Documento16 páginasBiorreactores Unidad 4David Hernandez Carlos100% (1)
- Etapas de La Reproducción HumanaDocumento2 páginasEtapas de La Reproducción HumanaCeballos Imelda100% (1)
- Glándula Timo-Aspectos Morfofuncionales y ClínicosDocumento14 páginasGlándula Timo-Aspectos Morfofuncionales y ClínicosRomero ErwinAún no hay calificaciones
- TimoDocumento4 páginasTimoAS GOAún no hay calificaciones
- Timo 140224184633 Phpapp01Documento29 páginasTimo 140224184633 Phpapp01Reinaldo FerreiraAún no hay calificaciones
- Taller Evaluativo Tejido Linfoide 2da ParteDocumento9 páginasTaller Evaluativo Tejido Linfoide 2da Partemauricio david sierra gonzalezAún no hay calificaciones
- Taller Evaluativo Tejido Linfoide 2da ParteDocumento8 páginasTaller Evaluativo Tejido Linfoide 2da Partemauricio david sierra gonzalez0% (1)
- Organos LinfoidesDocumento20 páginasOrganos Linfoidesmiguel angel100% (13)
- Proyecto de Ciencias Biologia. BachilleratoDocumento14 páginasProyecto de Ciencias Biologia. BachilleratoVictoria MijaresAún no hay calificaciones
- Timo Prueba 2Documento5 páginasTimo Prueba 2Eduardo De YcazaAún no hay calificaciones
- Organos Linfoides Primarios y SegundariosDocumento12 páginasOrganos Linfoides Primarios y SegundariosKaira Zonard100% (2)
- TimoDocumento4 páginasTimoDanny González CuetoAún no hay calificaciones
- TIMODocumento18 páginasTIMOangeliita1407Aún no hay calificaciones
- EMBRIOLOGIADocumento8 páginasEMBRIOLOGIAKarla Juarez100% (1)
- Orien 15Documento24 páginasOrien 15Rosdairy BorjaAún no hay calificaciones
- Timo 1Documento8 páginasTimo 1julio espinozaAún no hay calificaciones
- 07 Timo - Componentes Celulares 2019Documento2 páginas07 Timo - Componentes Celulares 2019Mariano QuirozAún no hay calificaciones
- Timo ExpoDocumento12 páginasTimo Expogarcialiconadianalaura4im12Aún no hay calificaciones
- El Timo MorofofisiologiaDocumento7 páginasEl Timo MorofofisiologiaMarco RojasAún no hay calificaciones
- Glandula Timo.Documento4 páginasGlandula Timo.María Fernanda PossoAún no hay calificaciones
- TIMODocumento4 páginasTIMOMaria Camila González100% (1)
- Sistema Endocrino)Documento7 páginasSistema Endocrino)Dainara ReyesAún no hay calificaciones
- Bazo y TimoDocumento7 páginasBazo y TimoBradem Maquera HuamaniAún no hay calificaciones
- Histología Del TimoDocumento3 páginasHistología Del TimoSandra Romero100% (1)
- El Timo y El BazoDocumento14 páginasEl Timo y El BazoYeseniaAún no hay calificaciones
- ESPERMATOGENESISDocumento15 páginasESPERMATOGENESISMiriam E. MairimAún no hay calificaciones
- Organos y Tejidos Del Sistema InmuneDocumento6 páginasOrganos y Tejidos Del Sistema InmuneChristian GavilánezAún no hay calificaciones
- Embriologia Falta CaratulaDocumento40 páginasEmbriologia Falta CaratulaSaira Diaz AldanaAún no hay calificaciones
- Órganos LinfáticosDocumento11 páginasÓrganos LinfáticosANGELA CONSTANZA ROJASAún no hay calificaciones
- LINFOCITOS T. Octavio ReyesDocumento49 páginasLINFOCITOS T. Octavio ReyesMELISSA JOHANA GONZALEZ SALCEDOAún no hay calificaciones
- 02 - Resumen y Transcripcion - Organos y Celulas Del Tejido LinfoideDocumento9 páginas02 - Resumen y Transcripcion - Organos y Celulas Del Tejido LinfoideLuis David Bello RodríguezAún no hay calificaciones
- Procesos de Division CelularDocumento32 páginasProcesos de Division CelularDiego FernandoAún no hay calificaciones
- Seri Linfohematopoyetico 2022Documento17 páginasSeri Linfohematopoyetico 2022Jonathan CorreaAún no hay calificaciones
- Gametogenesis Espermatogenesisi Ovogenesis InformeDocumento15 páginasGametogenesis Espermatogenesisi Ovogenesis InformeBAUTISTA TERRONES GILMERAún no hay calificaciones
- G3s2imuno Células y Órganos Del Sistema InmuneDocumento6 páginasG3s2imuno Células y Órganos Del Sistema Inmunelucia mayancelaAún no hay calificaciones
- Organos y Tejidos Implicados en La Respuesta InmuneDocumento8 páginasOrganos y Tejidos Implicados en La Respuesta Inmunelezc239Aún no hay calificaciones
- Sistema HematopoyeticoDocumento19 páginasSistema HematopoyeticoAngélica MartinezAún no hay calificaciones
- Timo y Educacion Timica, TareaDocumento4 páginasTimo y Educacion Timica, TareaMauricio IríasAún no hay calificaciones
- TIMO (Hematopoyetico y Linfoideo ) - Morfologia 2010Documento9 páginasTIMO (Hematopoyetico y Linfoideo ) - Morfologia 2010LUZ_LATINAAún no hay calificaciones
- INMUNIDADDocumento21 páginasINMUNIDADLiz ValenzuelaAún no hay calificaciones
- TimoDocumento12 páginasTimoXimena AnguloAún no hay calificaciones
- Avances en El Conocimiento de La Espermatogénesis HumanaDocumento14 páginasAvances en El Conocimiento de La Espermatogénesis HumanaJenniffer RonquilloAún no hay calificaciones
- Hematopoyesis - Organos HematopoyeticosDocumento38 páginasHematopoyesis - Organos HematopoyeticosSTEPHANNY NICOLE SANCHEZ AUCCAPUMAAún no hay calificaciones
- Plan de Mejoramiento Biologia SeptimoDocumento4 páginasPlan de Mejoramiento Biologia SeptimoJosé Nicolás Díaz VelásquezAún no hay calificaciones
- Cuestionario Histo Uca 1Documento59 páginasCuestionario Histo Uca 1By JAIROAún no hay calificaciones
- TEMA 2. Células y Órganos Del Sistema Inmune y Circulación LinfocitariaDocumento22 páginasTEMA 2. Células y Órganos Del Sistema Inmune y Circulación LinfocitariaNina Souto BlancoAún no hay calificaciones
- Embriologia GeneralDocumento13 páginasEmbriologia Generalesk hAún no hay calificaciones
- Resumen Celulas Del Sistema InmuneDocumento2 páginasResumen Celulas Del Sistema InmuneCarlos SorianoAún no hay calificaciones
- Órganos LinfoidesDocumento9 páginasÓrganos LinfoidesLucia EscobosaAún no hay calificaciones
- Biología y Tecnolohía de La Reproducción I El Cuerpo Lúteo. Histología y FisiopatologíaDocumento9 páginasBiología y Tecnolohía de La Reproducción I El Cuerpo Lúteo. Histología y Fisiopatologíaangel chavezAún no hay calificaciones
- The Key Role of Vitamin A in Spermatogenesis (ESPAÑOL)Documento8 páginasThe Key Role of Vitamin A in Spermatogenesis (ESPAÑOL)oliverdelgado323Aún no hay calificaciones
- Órganos Linfoides PrimariosDocumento17 páginasÓrganos Linfoides PrimariosCARLOS DIEGO VILA MADRIDAún no hay calificaciones
- Práctica 1 Morfo 2021Documento8 páginasPráctica 1 Morfo 2021lyaAún no hay calificaciones
- Daño PlacentarioDocumento20 páginasDaño PlacentarioEdwin Luis Quispe RetamozoAún no hay calificaciones
- MeiosisDocumento12 páginasMeiosiselizabeth.lee2179Aún no hay calificaciones
- Hematopoyesis AAyFADocumento15 páginasHematopoyesis AAyFAmarytayupegAún no hay calificaciones
- Tema31 Reproductor MasculinoDocumento9 páginasTema31 Reproductor Masculinomoises chino bandaAún no hay calificaciones
- Qué Es La Médula ÓseaDocumento6 páginasQué Es La Médula ÓseaCamila AilenAún no hay calificaciones
- 02 Celulas, Moléculas, Tejidos y Circ Linf, Texto y FigurasDocumento14 páginas02 Celulas, Moléculas, Tejidos y Circ Linf, Texto y FigurasJohn FósilAún no hay calificaciones
- Bio017 Diana Martinez Seccion A4 (No Borrar)Documento12 páginasBio017 Diana Martinez Seccion A4 (No Borrar)Adrianni Martinez valdezAún no hay calificaciones
- Mecanismos de de DefensaDocumento15 páginasMecanismos de de DefensaARLEIDYS RODRÍGUEZ RODRÍGUEZAún no hay calificaciones
- Anemia Para Todos: Medicina Para TodosDe EverandAnemia Para Todos: Medicina Para TodosCalificación: 3 de 5 estrellas3/5 (1)
- Estudios Sobre La Parte Especial Del Derecho Penal ChilenoDocumento383 páginasEstudios Sobre La Parte Especial Del Derecho Penal ChilenoM100% (1)
- Manual Casos PrácticosDocumento1 páginaManual Casos PrácticosMAún no hay calificaciones
- Nueva Constitución y Pueblos IndígenasDocumento205 páginasNueva Constitución y Pueblos IndígenasKRNPAZ100% (3)
- !… Escucha, Winka… ! Cuatro Ensayos de Historia Nacional Mapuche y Un Epílogo Sobre El FuturoDocumento1 página!… Escucha, Winka… ! Cuatro Ensayos de Historia Nacional Mapuche y Un Epílogo Sobre El FuturoRomero Mora-Caimanque AguirreAún no hay calificaciones
- Manejo Sustentable Bosque NativoDocumento1 páginaManejo Sustentable Bosque NativoMAún no hay calificaciones
- Fallo de La Corte Suprema PDFDocumento12 páginasFallo de La Corte Suprema PDFMAún no hay calificaciones
- Recomendaciones Al Estado Chileno Sobre Derechos HumanosDocumento312 páginasRecomendaciones Al Estado Chileno Sobre Derechos HumanosMAún no hay calificaciones
- Comision Pueblo MapucheDocumento4 páginasComision Pueblo MapucheMAún no hay calificaciones
- Bobbio Unidad 1Documento8 páginasBobbio Unidad 1Fabio JuarezAún no hay calificaciones
- Informe Reconstruccion Post TerremotoDocumento98 páginasInforme Reconstruccion Post TerremotoMAún no hay calificaciones
- Recomendaciones Al Estado Chileno Sobre Derechos HumanosDocumento312 páginasRecomendaciones Al Estado Chileno Sobre Derechos HumanosMAún no hay calificaciones
- Ley Sobre Recuperación Del Bosque Nativo y Fomento Forestal N°20.283Documento92 páginasLey Sobre Recuperación Del Bosque Nativo y Fomento Forestal N°20.283Enzo MartelliAún no hay calificaciones
- Ley Sobre Recuperación Del Bosque Nativo y Fomento Forestal N°20.283Documento92 páginasLey Sobre Recuperación Del Bosque Nativo y Fomento Forestal N°20.283Enzo MartelliAún no hay calificaciones
- Informe Sobre TPP ChileDocumento56 páginasInforme Sobre TPP ChileMAún no hay calificaciones
- Perspectiva de GéneroDocumento7 páginasPerspectiva de GéneroMAún no hay calificaciones
- Juicio de Roma Al Plan CondorDocumento36 páginasJuicio de Roma Al Plan CondorMAún no hay calificaciones
- REFLEXIONESDocumento14 páginasREFLEXIONESMAún no hay calificaciones
- Oscar Oszlak - Formación Histórica Del Estado Argentino: La Conquista Del OrdenDocumento27 páginasOscar Oszlak - Formación Histórica Del Estado Argentino: La Conquista Del OrdenMateo Nielsen Obieta100% (1)
- Material Estudio Intro Estudio CA Politica CPyRRIIDocumento127 páginasMaterial Estudio Intro Estudio CA Politica CPyRRIIElkin VelasteguíAún no hay calificaciones
- Sociología 1 2019Documento2 páginasSociología 1 2019MAún no hay calificaciones
- Informe Sobre TPP ChileDocumento56 páginasInforme Sobre TPP ChileMAún no hay calificaciones
- AasmxsaDocumento1 páginaAasmxsaMAún no hay calificaciones
- Derecho IndiceDocumento3 páginasDerecho IndiceMAún no hay calificaciones
- Curriculum Vitae Carlos Ramirez NOVIEMBRE 2014Documento6 páginasCurriculum Vitae Carlos Ramirez NOVIEMBRE 2014Preuniversitario KayrosAún no hay calificaciones
- Musgo 2Documento4 páginasMusgo 2karen taleroAún no hay calificaciones
- FisiologíaDocumento28 páginasFisiologíaF.C. BorrásAún no hay calificaciones
- Historia de La MicrobiologíaDocumento4 páginasHistoria de La MicrobiologíaKarina ContrerasAún no hay calificaciones
- BioquímicaDocumento36 páginasBioquímicaRosa JaimesAún no hay calificaciones
- Lesion Celular PatologíaDocumento29 páginasLesion Celular PatologíaSilvia Fernanda Rojas SerranoAún no hay calificaciones
- Averts PDFDocumento1 páginaAverts PDFAlejandro RamirezAún no hay calificaciones
- Clase+1 Introducción+Conservación+Biodiversidad PDFDocumento90 páginasClase+1 Introducción+Conservación+Biodiversidad PDFGeraldine Catalán CáceresAún no hay calificaciones
- Clasificaciones Historicas de La Raza HumanaDocumento16 páginasClasificaciones Historicas de La Raza HumanaJosé Paredes DiazAún no hay calificaciones
- Tema 8 Sistema de Endomembranas Reticulo EndoplasmaticoDocumento16 páginasTema 8 Sistema de Endomembranas Reticulo EndoplasmaticoValentina CelaniAún no hay calificaciones
- Historia de La PsicopedagogíaDocumento72 páginasHistoria de La PsicopedagogíaanaAún no hay calificaciones
- ADN - Nivel: SecundariaDocumento2 páginasADN - Nivel: SecundariaAnaRopeAún no hay calificaciones
- PSYCH-K LibroDocumento91 páginasPSYCH-K LibroJeronimo MillanAún no hay calificaciones
- El Control Naturalde Insectosenel Ecosistema Cafetero ColombianoDocumento222 páginasEl Control Naturalde Insectosenel Ecosistema Cafetero ColombianoJORGE IVAN LOAIZA BONILLAAún no hay calificaciones
- DifteriaDocumento10 páginasDifteriaDaryRangelAún no hay calificaciones
- Mononuclosis InfecciosaDocumento12 páginasMononuclosis InfecciosaIvhett MJ100% (1)
- Clase 9 - Reacciones Del Ciclo de Krebs y Balance Energético, Mecanismos de Regulación. Interacción Con Otras Rutas Metabólicas, Ciclo Del GlioxilatoDocumento29 páginasClase 9 - Reacciones Del Ciclo de Krebs y Balance Energético, Mecanismos de Regulación. Interacción Con Otras Rutas Metabólicas, Ciclo Del GlioxilatoAlex Ostos AlvaAún no hay calificaciones
- Interacción AlostéricaDocumento3 páginasInteracción AlostéricaNathaly Rojas Jaramillo100% (1)
- Dónde Se Produce La GlucólisisDocumento2 páginasDónde Se Produce La GlucólisisPabloGordón100% (1)
- Articulo Cientifico - Parasitosis - AmebiasisDocumento11 páginasArticulo Cientifico - Parasitosis - AmebiasisWaltr Q CastrellonAún no hay calificaciones
- Pci Final 2018-2022 Uets PDFDocumento620 páginasPci Final 2018-2022 Uets PDFLuis Antonio Morales PullasAún no hay calificaciones
- Sanidad Animal Ganaderia EcoDocumento6 páginasSanidad Animal Ganaderia EcoAna M GómezAún no hay calificaciones
- Parasitología Medica GeneralidadesDocumento44 páginasParasitología Medica GeneralidadesElisa Chinchilla100% (1)
- Un 1plan de Clase Naturales 8Documento14 páginasUn 1plan de Clase Naturales 8mprm2579Aún no hay calificaciones
- LeflunomidaDocumento3 páginasLeflunomidaregulohAún no hay calificaciones
- Clase de MutagenesisDocumento18 páginasClase de MutagenesisPero CómoAún no hay calificaciones
- Manual de Procedimientos Tecnicos para El Diagnostico Micologico - FinalDocumento139 páginasManual de Procedimientos Tecnicos para El Diagnostico Micologico - FinalIgnacio Candia Ramos100% (1)
- Tarea 1 BiologiaDocumento3 páginasTarea 1 BiologiaEnrique Lezcano (C.E courier express)Aún no hay calificaciones