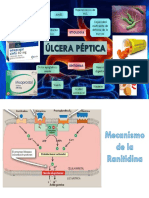
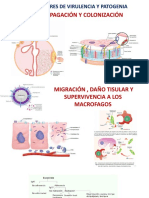
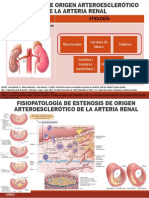
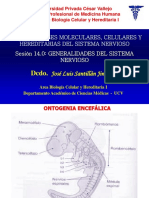
También podría gustarte
- Uniones CelularesDocumento6 páginasUniones CelularesMaria Melissa Tapias PerezAún no hay calificaciones
- Uniones IntercelularesDocumento15 páginasUniones IntercelularesVERA SILVANA GARAMENDI SACSARAAún no hay calificaciones
- Objetivos Uniones Intercelulares Adhesion Celular y Matriz ExtracelularDocumento9 páginasObjetivos Uniones Intercelulares Adhesion Celular y Matriz Extracelularapi-3696656100% (12)
- Adherencia Celular en Las Membranas EpitelialesDocumento7 páginasAdherencia Celular en Las Membranas EpitelialesCarlos Alfonso Catuto HerasAún no hay calificaciones
- Complejos de UniónDocumento7 páginasComplejos de UniónAnalia Moraes100% (1)
- Uniones IntercelularesDocumento6 páginasUniones IntercelularesAgustina NuñezAún no hay calificaciones
- R 17Documento9 páginasR 17danitzaAún no hay calificaciones
- HISTOLOGIADocumento4 páginasHISTOLOGIAdavidAún no hay calificaciones
- Adhesión y Unión CelularDocumento5 páginasAdhesión y Unión CelularFrancisco AriasAún no hay calificaciones
- Inter Acci OnesDocumento19 páginasInter Acci OnesErnesto Batista GarcíaAún no hay calificaciones
- Union CelularDocumento11 páginasUnion CelularHEBER LOPEZ GALDOSAún no hay calificaciones
- Uniones IntercelularesDocumento4 páginasUniones IntercelularesAlexiiz AntiñancoAún no hay calificaciones
- Uniones IntercelularesDocumento2 páginasUniones IntercelularesJavier Rama GonzálezAún no hay calificaciones
- Guia Adhesion Celular - 4toELECTIVODocumento4 páginasGuia Adhesion Celular - 4toELECTIVOMaria Alicia Bravo ArriagadaAún no hay calificaciones
- Adhesión y Uniones CelularesDocumento4 páginasAdhesión y Uniones Celularespimenteltamara2Aún no hay calificaciones
- Uniones CelularesDocumento3 páginasUniones CelularesAndreaAún no hay calificaciones
- Lectura Semana 1Documento7 páginasLectura Semana 1Study zoneAún no hay calificaciones
- Biologia Celular - Clase 05Documento58 páginasBiologia Celular - Clase 05api-3701781Aún no hay calificaciones
- Tema 4. Uniones IntercelularesDocumento4 páginasTema 4. Uniones IntercelularesMercedes MartínAún no hay calificaciones
- Uniones CelularesDocumento7 páginasUniones CelularesLeninLeo 18Aún no hay calificaciones
- HistologiaDocumento11 páginasHistologiaBianca BravoAún no hay calificaciones
- Uniones AdherentesDocumento34 páginasUniones Adherentesnoge77Aún no hay calificaciones
- Uniones CelularesDocumento3 páginasUniones Celularesalejandroguerrero1711Aún no hay calificaciones
- Adhesión CelularDocumento1 páginaAdhesión CelularTiare VegaAún no hay calificaciones
- Uniones de OclusiónDocumento12 páginasUniones de OclusiónAnonymous 9xk31PFOznAún no hay calificaciones
- Uniones Ceulares 2022Documento20 páginasUniones Ceulares 2022Fran TejadaAún no hay calificaciones
- Actividad 11 TallerSeñalizacionCelular-InteracciónCélulaCélulaDocumento9 páginasActividad 11 TallerSeñalizacionCelular-InteracciónCélulaCélulaAndrés FelipeAún no hay calificaciones
- TemaDocumento8 páginasTemaJoseAún no hay calificaciones
- Tema11: Uniones IntercelularesDocumento6 páginasTema11: Uniones IntercelularesTBProtein100% (10)
- Uniones CelularesDocumento9 páginasUniones CelularesFranklin Clark0% (1)
- PDF Las Interacciones de Las Celulas Entre Si Grupo 5 DDDocumento7 páginasPDF Las Interacciones de Las Celulas Entre Si Grupo 5 DDpilar morochoAún no hay calificaciones
- Seminario Matriz Extracelular y Unión CelularDocumento16 páginasSeminario Matriz Extracelular y Unión CelularNoemi EscobarAún no hay calificaciones
- Especializaciones de La Región Lateral - JONALISDocumento2 páginasEspecializaciones de La Región Lateral - JONALISMichael GonzálezAún no hay calificaciones
- TallerSeñalizacionCelular InteracciónCélulaCélulaDocumento10 páginasTallerSeñalizacionCelular InteracciónCélulaCélulaJesus Martinez0% (1)
- APUNTESDocumento24 páginasAPUNTESJose M. Gutierrez JimenezAún no hay calificaciones
- Uniones CelularesDocumento14 páginasUniones CelularesGabriela EscalanteAún no hay calificaciones
- Parte Alan Daniel Cortez Cruz 30543Documento3 páginasParte Alan Daniel Cortez Cruz 30543Cortez Sebastian AlanAún no hay calificaciones
- Uniones CelularesDocumento6 páginasUniones CelularesMilagros RuferAún no hay calificaciones
- Uniones IntercellDocumento5 páginasUniones Intercellgustavoarrieta59Aún no hay calificaciones
- Matriz Extracelular de BiologíaDocumento3 páginasMatriz Extracelular de BiologíamariaoaAún no hay calificaciones
- Cara LateralDocumento14 páginasCara LateralIdaLii SancheezAún no hay calificaciones
- Resumen Principios de Fisiologia y AnatomiaDocumento2 páginasResumen Principios de Fisiologia y AnatomiaMiguel Angel Gonzalez VargasAún no hay calificaciones
- 4° Medio Prueba Adhesion CelularDocumento4 páginas4° Medio Prueba Adhesion CelularJenny Castro Jara0% (2)
- T.5 Uniones y Adhesiones CelularesDocumento9 páginasT.5 Uniones y Adhesiones CelularesNatalia Villar CrespoAún no hay calificaciones
- Tipos de Uniones CelularesDocumento2 páginasTipos de Uniones Celularesdanna celisAún no hay calificaciones
- Adhesión CelularDocumento33 páginasAdhesión CelularArmando BravoAún no hay calificaciones
- Unión CelularDocumento10 páginasUnión CelularAuRee' DenizAún no hay calificaciones
- Uniones CelularesDocumento22 páginasUniones Celularesjulietttta100% (1)
- S2 - Tejido EpitelialDocumento4 páginasS2 - Tejido EpitelialCandy Campos QuispeAún no hay calificaciones
- Guia Adhesion Celular 2011Documento4 páginasGuia Adhesion Celular 2011Clarys DiazAún no hay calificaciones
- Matrix ExtracellDocumento7 páginasMatrix Extracellgustavoarrieta59Aún no hay calificaciones
- La Adhesión en La Membrana CelularDocumento6 páginasLa Adhesión en La Membrana Celularjulian.montes.95julianAún no hay calificaciones
- Guia Adhesion Celular 2017Documento4 páginasGuia Adhesion Celular 2017Carolina Severino0% (1)
- Las Células y Su Contexto SocialDocumento27 páginasLas Células y Su Contexto SocialRocio Rosas LópezAún no hay calificaciones
- Clase - 7 Adhesión Celular-4 Dif.Documento28 páginasClase - 7 Adhesión Celular-4 Dif.KAREN VIVIANA PAVEZ RODRIGUEZAún no hay calificaciones
- Diferenciaciones y MatrizDocumento17 páginasDiferenciaciones y MatrizmarinainesgonAún no hay calificaciones
- BC 10 NecroticaenfmedDocumento4 páginasBC 10 NecroticaenfmedCasa LimpiezaAún no hay calificaciones
- Uniones Celulares, MEC y MBDocumento4 páginasUniones Celulares, MEC y MBFabiana AvalosAún no hay calificaciones
- Introducción a la Biología: RESÚMENES UNIVERSITARIOSDe EverandIntroducción a la Biología: RESÚMENES UNIVERSITARIOSCalificación: 5 de 5 estrellas5/5 (1)
- Guía de Prácticas #05 PDFDocumento5 páginasGuía de Prácticas #05 PDFVictor Pier Chuquipoma RojasAún no hay calificaciones
- Insuficiencia Respiratoria: Definición EtiologíaDocumento4 páginasInsuficiencia Respiratoria: Definición EtiologíaVictor Pier Chuquipoma RojasAún no hay calificaciones
- Caso ClicnicoDocumento18 páginasCaso ClicnicoVictor Pier Chuquipoma RojasAún no hay calificaciones
- Semana 1Documento2 páginasSemana 1Victor Pier Chuquipoma RojasAún no hay calificaciones
- Semana 1Documento2 páginasSemana 1Victor Pier Chuquipoma RojasAún no hay calificaciones
- Funda ExpoDocumento4 páginasFunda ExpoVictor Pier Chuquipoma RojasAún no hay calificaciones
- PPTDocumento2 páginasPPTVictor Pier Chuquipoma RojasAún no hay calificaciones
- Biosíntesis de Estrogenos BioloDocumento2 páginasBiosíntesis de Estrogenos BioloVictor Pier Chuquipoma RojasAún no hay calificaciones
- 002 MAIS - Principios Integradores 2017-2Documento27 páginas002 MAIS - Principios Integradores 2017-2Victor Pier Chuquipoma RojasAún no hay calificaciones
- Churcampa AvancesDocumento17 páginasChurcampa AvancesVictor Pier Chuquipoma RojasAún no hay calificaciones
- Glándula Tiroides y ParatiroidesDocumento5 páginasGlándula Tiroides y ParatiroidesVictor Pier Chuquipoma RojasAún no hay calificaciones
- Analisis Del Centro de Salud Victor LarcoDocumento60 páginasAnalisis Del Centro de Salud Victor LarcoVictor Pier Chuquipoma Rojas50% (2)
- Silabo - Salud PúblicaDocumento11 páginasSilabo - Salud PúblicaVictor Pier Chuquipoma RojasAún no hay calificaciones
- RM 204 2015 MINSA Ficha Familiar.Documento7 páginasRM 204 2015 MINSA Ficha Familiar.susanAún no hay calificaciones
- SOCIODEMOGRAFIADocumento8 páginasSOCIODEMOGRAFIAitmi4Aún no hay calificaciones
- Tipos de Piramides PDFDocumento8 páginasTipos de Piramides PDFpablovillar76Aún no hay calificaciones
- Descentralizacion y Gestion TerritorialDocumento14 páginasDescentralizacion y Gestion TerritorialVictor Pier Chuquipoma RojasAún no hay calificaciones
- Diapositivas Redaccion UniversitariasDocumento17 páginasDiapositivas Redaccion UniversitariasVictor Pier Chuquipoma RojasAún no hay calificaciones
- Sistema Digestivo Clase 1 Dr. BarónDocumento38 páginasSistema Digestivo Clase 1 Dr. BarónVictor Pier Chuquipoma RojasAún no hay calificaciones
- Gu A Taller Instrumentos de FamiliaDocumento13 páginasGu A Taller Instrumentos de FamiliaFurboo Toretto ValdezAún no hay calificaciones
- FamiliaDocumento6 páginasFamiliaCaro De Lima MalladaAún no hay calificaciones
- Semana 1Documento2 páginasSemana 1Victor Pier Chuquipoma RojasAún no hay calificaciones
- 5336material Profesores UD 1 - Funciones y RolesDocumento23 páginas5336material Profesores UD 1 - Funciones y RolesVictor Pier Chuquipoma RojasAún no hay calificaciones
- Sistema Digestivo Clase 3 Dr. BarónDocumento44 páginasSistema Digestivo Clase 3 Dr. BarónVictor Pier Chuquipoma RojasAún no hay calificaciones
- Mecanismo Meiosis Clase 8 Dr. BarónDocumento41 páginasMecanismo Meiosis Clase 8 Dr. BarónVictor Pier Chuquipoma RojasAún no hay calificaciones
- Generalidades Sistema NerviosoDocumento34 páginasGeneralidades Sistema NerviosoVictor Pier Chuquipoma RojasAún no hay calificaciones
- Sistema Urinario Clase 5 Dr. BarónDocumento37 páginasSistema Urinario Clase 5 Dr. BarónVictor Pier Chuquipoma RojasAún no hay calificaciones
- Sistema Urinario Clase 6 Dr. BarónDocumento34 páginasSistema Urinario Clase 6 Dr. BarónVictor Pier Chuquipoma RojasAún no hay calificaciones
- Duplicción Transcripción Código Genetico TraducciónDocumento19 páginasDuplicción Transcripción Código Genetico TraducciónVictor Pier Chuquipoma RojasAún no hay calificaciones
- Meiosis I IIDocumento22 páginasMeiosis I IIVictor Pier Chuquipoma RojasAún no hay calificaciones
- Estructura BacteriasDocumento4 páginasEstructura Bacteriasjessica perezAún no hay calificaciones
- Epigenetic Molecular Mechanisms in InsectsDocumento28 páginasEpigenetic Molecular Mechanisms in InsectsZaidy OlayaAún no hay calificaciones
- Moleculas RepresorasDocumento44 páginasMoleculas RepresorasGioborjAún no hay calificaciones
- Introduccion para El 1Documento4 páginasIntroduccion para El 1Elianne Alexandra Diaz SaldañaAún no hay calificaciones
- Wuolah Free TEMA 9 Acoplamiento Excitacion Contraccion PDFDocumento9 páginasWuolah Free TEMA 9 Acoplamiento Excitacion Contraccion PDFZaira Sánchez GonzaloAún no hay calificaciones
- CoenzimasDocumento8 páginasCoenzimasCIELOAún no hay calificaciones
- Cuestionario 8 y 9 BioquímicaDocumento3 páginasCuestionario 8 y 9 BioquímicaNick Soto FlukerAún no hay calificaciones
- Pre Informe Interm. MetabolicosDocumento5 páginasPre Informe Interm. MetabolicosAna GonzálezAún no hay calificaciones
- Bases Moleculares de La MutacionDocumento55 páginasBases Moleculares de La MutacionDiana GonzalezAún no hay calificaciones
- Biolog÷a-Examen 1-2012Documento2 páginasBiolog÷a-Examen 1-2012ConchaAún no hay calificaciones
- CUESTIONARIODocumento4 páginasCUESTIONARIOElias MaytaAún no hay calificaciones
- Inmunologia (1) Lizzzz - OdtDocumento10 páginasInmunologia (1) Lizzzz - Odtwilber carvajalAún no hay calificaciones
- Universidad Del Cauca Facultad de CienciDocumento7 páginasUniversidad Del Cauca Facultad de CienciDani Anto Del VaAún no hay calificaciones
- Importancia de Los Marcadores Moleculares para Estudio de Diversidad Genetica Vegetal.Documento16 páginasImportancia de Los Marcadores Moleculares para Estudio de Diversidad Genetica Vegetal.ROBINSON ALEJANDRO PEREZ MANOSALVA100% (1)
- ANEXOSDocumento6 páginasANEXOSJacqueline CLAún no hay calificaciones
- Enfermedades Por Alteración GenéticaDocumento10 páginasEnfermedades Por Alteración GenéticaOriana Victoria Osorio SpissoAún no hay calificaciones
- BiologiaDocumento3 páginasBiologiaJOSE .CAún no hay calificaciones
- Cinetica Enzimatica ReporteDocumento6 páginasCinetica Enzimatica ReporteGabriel LVAún no hay calificaciones
- Seminario Tema: PCR (Reacción en Cadena de La Polimerasa)Documento3 páginasSeminario Tema: PCR (Reacción en Cadena de La Polimerasa)Claudia PaezAún no hay calificaciones
- Extraccion Adn PlasmidicoDocumento5 páginasExtraccion Adn PlasmidicoAlejandro FrancoAún no hay calificaciones
- Comportamiento de La Membrana CelularDocumento14 páginasComportamiento de La Membrana CelularemmanuelAún no hay calificaciones
- Práctica de Identificación de Aminoácidos y ProteínasDocumento9 páginasPráctica de Identificación de Aminoácidos y ProteínasDayana Guadalupe Medina MendozaAún no hay calificaciones
- Luz AzulDocumento2 páginasLuz AzulCecilia NajeraAún no hay calificaciones
- Introduccion A La FarmacologiaDocumento117 páginasIntroduccion A La Farmacologiajosefina diaz100% (1)
- Cap. 16 Señalización Celular (PARTE 1)Documento37 páginasCap. 16 Señalización Celular (PARTE 1)FernanBoykaRodriguezValenzuelaAún no hay calificaciones
- Enzimas 10Documento15 páginasEnzimas 10Maria Victoria CaballeroAún no hay calificaciones
- Cap 74 - Introducion A La EndocrinologiaDocumento2 páginasCap 74 - Introducion A La EndocrinologiaDARYL DAFNE ORTIZ REYESAún no hay calificaciones
- Replicacion de AdnDocumento13 páginasReplicacion de AdnYaritza Topete GonzalezAún no hay calificaciones
- Resumen LabDocumento18 páginasResumen LabKaren VanesaAún no hay calificaciones
- Tema 5. Obtención y Producción de AnticuerposDocumento13 páginasTema 5. Obtención y Producción de AnticuerposSofía Parra ScafiezzoAún no hay calificaciones