También podría gustarte
- Práctica 17. NucleoloDocumento5 páginasPráctica 17. NucleoloAndrea TovarAún no hay calificaciones
- Tarea Resumen El Nucleolo Como Un Regulador Del Envejecimiento CelularDocumento4 páginasTarea Resumen El Nucleolo Como Un Regulador Del Envejecimiento Celularkatherin PCAún no hay calificaciones
- Taller Sobre Lectura Crítica - Ana Milena RuizDocumento4 páginasTaller Sobre Lectura Crítica - Ana Milena RuizAna Milena Ruiz RomeroAún no hay calificaciones
- Nucleo y NucleoloDocumento3 páginasNucleo y NucleoloFelix Gutierrez100% (1)
- NucleologenesisDocumento48 páginasNucleologenesisRodolfo Castillo50% (2)
- Informe de Exposición NúcleoDocumento14 páginasInforme de Exposición Núcleomafer salazarAún no hay calificaciones
- INTRODUCCION3333Documento2 páginasINTRODUCCION3333JorgeAún no hay calificaciones
- Nucleo y NucleoloDocumento3 páginasNucleo y NucleoloJuan Carlos Vázquez92% (13)
- Estructura y función del nucléolo celular enDocumento29 páginasEstructura y función del nucléolo celular enFredy Alberto CabasAún no hay calificaciones
- Arquitectura nuclear dinámicaDocumento9 páginasArquitectura nuclear dinámicaJhon RVAún no hay calificaciones
- Informe de BioCel - NucleoloDocumento7 páginasInforme de BioCel - NucleoloJuan Llontop SolanoAún no hay calificaciones
- El Nucleolo en EspanolDocumento7 páginasEl Nucleolo en Espanolmateo charrisAún no hay calificaciones
- Nucleo ArquitecturaDocumento8 páginasNucleo Arquitecturaarcadia_hbAún no hay calificaciones
- Unidad 9 ReplicDocumento29 páginasUnidad 9 ReplicLUDMILA FERNÁNDEZAún no hay calificaciones
- Unidad 09 ReplicacionDocumento12 páginasUnidad 09 ReplicacionMicaela FragaAún no hay calificaciones
- El Nucleolo (Resumen)Documento6 páginasEl Nucleolo (Resumen)Angel Maqquerhua PereyraAún no hay calificaciones
- Los límites entre la histología y la bioquímica: observando al núcleo celular con ojo histológico-bioquímicoDocumento16 páginasLos límites entre la histología y la bioquímica: observando al núcleo celular con ojo histológico-bioquímicoedson floresAún no hay calificaciones
- Articulos CientificosDocumento4 páginasArticulos CientificosAlejandra Arcos GuerreroAún no hay calificaciones
- El NucleoDocumento10 páginasEl NucleoSmith Rels100% (1)
- Nucleo InterfasticoDocumento18 páginasNucleo InterfasticoAnahi Alban100% (1)
- Trabajo Práctico #2.Documento23 páginasTrabajo Práctico #2.Juan Cruz FARFANAún no hay calificaciones
- Triptico Celula BiologiaDocumento3 páginasTriptico Celula BiologiaMIRNA75% (4)
- La Frontera Genetica Molecular de La EnfermedadDocumento526 páginasLa Frontera Genetica Molecular de La EnfermedadDeivy Horta Tkd0% (1)
- Info. Importante Nucleo y NucleoloDocumento4 páginasInfo. Importante Nucleo y NucleoloAlejandraAún no hay calificaciones
- Nucleo InterfasicoDocumento16 páginasNucleo InterfasicoVane PonsAún no hay calificaciones
- La Frontera Genetica - MolecularDocumento526 páginasLa Frontera Genetica - MolecularFrancis Vallecillo PadillaAún no hay calificaciones
- El NucleoloDocumento6 páginasEl NucleoloHanna ZavalaAún no hay calificaciones
- Biologia Trabajo CelulaDocumento49 páginasBiologia Trabajo CelulaKARLA BAUTISTAAún no hay calificaciones
- El Daño Tisular y La Senescencia Proporcionan Señales Críticas para La Reprogramación Celular in VivoDocumento24 páginasEl Daño Tisular y La Senescencia Proporcionan Señales Críticas para La Reprogramación Celular in VivoManuel David Fuentes GarciaAún no hay calificaciones
- NUCLEODocumento15 páginasNUCLEOEsmi Esmi TexotlticitlAún no hay calificaciones
- Núcleo CelularDocumento32 páginasNúcleo CelularAlisson PonceAún no hay calificaciones
- Clase Estructura Del NucleoDocumento90 páginasClase Estructura Del Nucleohghbkgykmgxyh100% (1)
- Genética SeminarioDocumento7 páginasGenética SeminarioMayerling V. RiosAún no hay calificaciones
- Resumen Nucleo EYFAMDocumento8 páginasResumen Nucleo EYFAMBbm NosbornAún no hay calificaciones
- Semana 7 - TeoriaDocumento23 páginasSemana 7 - TeoriajoselinAún no hay calificaciones
- Informe de Biologia Nucleo CelularDocumento11 páginasInforme de Biologia Nucleo CelularelmerAún no hay calificaciones
- Informe de Biologia Nucleo CelularDocumento11 páginasInforme de Biologia Nucleo CelularelmerAún no hay calificaciones
- El Núcleo CelularDocumento3 páginasEl Núcleo CelularGinger AvilesAún no hay calificaciones
- Guía TP 5Documento16 páginasGuía TP 5Dani Morales DellatorreAún no hay calificaciones
- Que Es El Jugo NuclearDocumento6 páginasQue Es El Jugo NuclearPaúl Fernando100% (1)
- Envuelta nuclear, el protector del núcleoDocumento20 páginasEnvuelta nuclear, el protector del núcleomanuel rodriguesAún no hay calificaciones
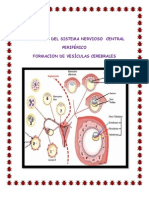
- Sistema Nervioso y Vesiculas CerebralesDocumento35 páginasSistema Nervioso y Vesiculas Cerebraleslorietarosemary100% (1)
- Nucleo CelularDocumento8 páginasNucleo CelularJHARIM NOELY AUCCAPIÑA GUILLENAún no hay calificaciones
- Organización NuclearDocumento6 páginasOrganización NuclearLuciana ColqueAún no hay calificaciones
- Núcleo CelularDocumento6 páginasNúcleo CelularBetzy Valery Poma PerezAún no hay calificaciones
- NucleoloDocumento2 páginasNucleoloMorales Tecalero Donaldo100% (1)
- Células del sistema nerviosoDocumento12 páginasCélulas del sistema nerviosoSamayraAún no hay calificaciones
- NucleoloDocumento15 páginasNucleololeAún no hay calificaciones
- Ciclo celular: fases y regulaciónDocumento2 páginasCiclo celular: fases y regulaciónAna Martínez TorresAún no hay calificaciones
- PANIAGUA Biologia Celular 3a Edicion - Estructura y Expresión Genética - ComprimidoDocumento59 páginasPANIAGUA Biologia Celular 3a Edicion - Estructura y Expresión Genética - ComprimidosuldenyAún no hay calificaciones
- Estructura y funciones del núcleo celularDocumento17 páginasEstructura y funciones del núcleo celularVanessa PeñaAún no hay calificaciones
- Tema 10_El núcleo_Reproducción celularDocumento23 páginasTema 10_El núcleo_Reproducción celularcandelaropero2006Aún no hay calificaciones
- Organelas Celulares: Núcleo y Ácidos NucleicosDocumento17 páginasOrganelas Celulares: Núcleo y Ácidos NucleicosMarcela BarriosAún no hay calificaciones
- Semana 7 Teoria - Nucleo CelularDocumento17 páginasSemana 7 Teoria - Nucleo CelularEliza huamaniAún no hay calificaciones
- Articulo Martinez Miguel JenniferDocumento47 páginasArticulo Martinez Miguel Jenniferkatherin PCAún no hay calificaciones
- Nucleo CelularDocumento3 páginasNucleo CelularMarleni RobayoAún no hay calificaciones
- Ultra Estructura Del Núcleo de La CélulaDocumento13 páginasUltra Estructura Del Núcleo de La CélulaEduardo Yamunaqué CastroAún no hay calificaciones
- Fundamentos y técnicas básicas en biología molecularDe EverandFundamentos y técnicas básicas en biología molecularAún no hay calificaciones
- Rejuvenecer Con El Plasma Sanguíneo De Los JóvenesDe EverandRejuvenecer Con El Plasma Sanguíneo De Los JóvenesCalificación: 5 de 5 estrellas5/5 (1)
- U1 Terorias Eticas 2020Documento22 páginasU1 Terorias Eticas 2020Randolfo MaximilianoAún no hay calificaciones
- U1 Acto Moral 2020Documento23 páginasU1 Acto Moral 2020Randolfo MaximilianoAún no hay calificaciones
- INTRODUCCION LOCOMOTOR Kine 2012Documento33 páginasINTRODUCCION LOCOMOTOR Kine 2012Randolfo MaximilianoAún no hay calificaciones
- Teoria de AproximacionesDocumento3 páginasTeoria de AproximacionesRandolfo MaximilianoAún no hay calificaciones
- Clase 1 Prof. SantosDocumento6 páginasClase 1 Prof. SantosRandolfo MaximilianoAún no hay calificaciones
- Actividad5cinebiotecnologa 120327014903 Phpapp02Documento10 páginasActividad5cinebiotecnologa 120327014903 Phpapp02Randolfo MaximilianoAún no hay calificaciones
- Teoria de AproximacionesDocumento3 páginasTeoria de AproximacionesRandolfo MaximilianoAún no hay calificaciones
- Curso Basico de BioeticaDocumento150 páginasCurso Basico de BioeticaRandolfo MaximilianoAún no hay calificaciones
- Virologia - Conceptos BasicosDocumento38 páginasVirologia - Conceptos BasicosRandolfo MaximilianoAún no hay calificaciones
- Relieve de Amer - Clase 5Documento2 páginasRelieve de Amer - Clase 5Randolfo MaximilianoAún no hay calificaciones
- Unidad 3 Taxonomia y Crecimiento Bacteriano PDFDocumento10 páginasUnidad 3 Taxonomia y Crecimiento Bacteriano PDFOctavioAún no hay calificaciones
- Material Teorico - FuerzaDocumento6 páginasMaterial Teorico - FuerzaRandolfo MaximilianoAún no hay calificaciones
- Apunte Acidos NucleicosDocumento16 páginasApunte Acidos NucleicosRandolfo MaximilianoAún no hay calificaciones
- Apunte Acidos NucleicosDocumento16 páginasApunte Acidos NucleicosRandolfo MaximilianoAún no hay calificaciones
- Teorico Practico 1 BioeticaDocumento36 páginasTeorico Practico 1 BioeticaRandolfo MaximilianoAún no hay calificaciones
- Acidos Nucleicos ResumenDocumento6 páginasAcidos Nucleicos ResumenRandolfo MaximilianoAún no hay calificaciones
- Bioetica 2019 AnalizarDocumento7 páginasBioetica 2019 AnalizarRandolfo MaximilianoAún no hay calificaciones
- Mapa Conceptual de Bacteriolog A y Virolog A M Dicas-5Documento2 páginasMapa Conceptual de Bacteriolog A y Virolog A M Dicas-5Randolfo MaximilianoAún no hay calificaciones
- El Nucléolo Como Un Regulador Del Envejecimiento CelularDocumento19 páginasEl Nucléolo Como Un Regulador Del Envejecimiento CelularRandolfo MaximilianoAún no hay calificaciones
- Guias de TPDocumento33 páginasGuias de TPRandolfo MaximilianoAún no hay calificaciones
- La Iglesia Naciã en La CalleDocumento1 páginaLa Iglesia Naciã en La CalleRandolfo MaximilianoAún no hay calificaciones
- Fermentación de AzucaresDocumento2 páginasFermentación de AzucaresRandolfo MaximilianoAún no hay calificaciones
- Estatuto Docente PDFDocumento144 páginasEstatuto Docente PDFRandolfo MaximilianoAún no hay calificaciones
- Adoracion Al Santisimo para Jovenes PDFDocumento10 páginasAdoracion Al Santisimo para Jovenes PDFJorge Gimenez100% (2)
- Guión Litúrgico de Solemne Eucaristía en Honor A San José (Recuperado Automáticamente)Documento2 páginasGuión Litúrgico de Solemne Eucaristía en Honor A San José (Recuperado Automáticamente)Randolfo Maximiliano100% (2)
- Guia de Actividades Ingenieria GeneticaDocumento9 páginasGuia de Actividades Ingenieria GeneticaRandolfo MaximilianoAún no hay calificaciones
- Manual Discapacidad 2019 BeneficiarioDocumento47 páginasManual Discapacidad 2019 BeneficiarioRandolfo MaximilianoAún no hay calificaciones
- Toma de MuestrasDocumento43 páginasToma de MuestrasRandolfo MaximilianoAún no hay calificaciones
- Evaluacion Compensatorio Febrero 2017Documento2 páginasEvaluacion Compensatorio Febrero 2017Randolfo MaximilianoAún no hay calificaciones
- Ingenieria GeneticaDocumento38 páginasIngenieria GeneticajavierAún no hay calificaciones
- P53Documento4 páginasP53CARLOS JESUS GARCIA HERNANDEZAún no hay calificaciones
- Regulación del tráfico de GPCR por arrestinasDocumento8 páginasRegulación del tráfico de GPCR por arrestinasWalter RamirezAún no hay calificaciones
- Tema 33 - Genes Supresores de Tumores SARADocumento10 páginasTema 33 - Genes Supresores de Tumores SARAJuan Manuel Morón OcañaAún no hay calificaciones
- p53 en El Proceso de Reparación de Daño de AdnDocumento5 páginasp53 en El Proceso de Reparación de Daño de AdnKaren GonzalesAún no hay calificaciones
- Radiación Ultravioleta Cancer Piel P53Documento8 páginasRadiación Ultravioleta Cancer Piel P53AlexRodrigoAún no hay calificaciones
- Receptores 7TMR arrestina tráfico señalizaciónDocumento13 páginasReceptores 7TMR arrestina tráfico señalizaciónSILVIA FERNANDA CASTILLO GOYENECHEAún no hay calificaciones
- Sarcomas RetroperitonealesDocumento58 páginasSarcomas RetroperitonealesSara MunozAún no hay calificaciones
- Cuerpos de CajalDocumento233 páginasCuerpos de CajalJuan Cruz FARFANAún no hay calificaciones
- Propiedades antígenos, anticuerpos e inmunidadDocumento95 páginasPropiedades antígenos, anticuerpos e inmunidaddannymalcacamposAún no hay calificaciones
- Tema11 - RESPUESTA CELULAR AL DAÑO DEL ADNDocumento34 páginasTema11 - RESPUESTA CELULAR AL DAÑO DEL ADNLidii monstersAún no hay calificaciones
- Boletín de La Academia de Medicina Chilena. 2007Documento246 páginasBoletín de La Academia de Medicina Chilena. 2007Camila Cisternas FloresAún no hay calificaciones
- Unidad 8 - NeoplasiasDocumento25 páginasUnidad 8 - NeoplasiasLourdes VelazquezAún no hay calificaciones
- Estabilización de Las Interacciones Proteína-Proteína en El Descubrimiento de FármacosDocumento57 páginasEstabilización de Las Interacciones Proteína-Proteína en El Descubrimiento de FármacosDavid VasquezAún no hay calificaciones
- Adenomas y Adenocarcinoma de GL HepatoidesDocumento32 páginasAdenomas y Adenocarcinoma de GL HepatoidesAlfredo MoralesAún no hay calificaciones
- p53 y su papel en el epitelio superficial del ovarioDocumento33 páginasp53 y su papel en el epitelio superficial del ovarioNestor lorenzoAún no hay calificaciones
- Osteosarcoma: A. ResumenDocumento33 páginasOsteosarcoma: A. ResumenHOSPITAL GENERAL DE LIBRESAún no hay calificaciones
- Bioinorgánica Del ZNDocumento10 páginasBioinorgánica Del ZNJesus27Aún no hay calificaciones
- Regulación Ciclo Celular y El CancerDocumento5 páginasRegulación Ciclo Celular y El CancerSantos Olav Godoy MoránAún no hay calificaciones
- Taller 1 y 2 Biologia Celular y MolecularDocumento53 páginasTaller 1 y 2 Biologia Celular y MolecularVanessa NúñezAún no hay calificaciones
- 1 Ciclo Celular y CáncerDocumento6 páginas1 Ciclo Celular y CáncerLISETH KATHERINE RAMIREZ GARCIAAún no hay calificaciones
- PICHER - El Nucléolo Como Sensor de Estrés Oxidativo. Implicaciones en Patologías Progeroides.Documento54 páginasPICHER - El Nucléolo Como Sensor de Estrés Oxidativo. Implicaciones en Patologías Progeroides.katherin PCAún no hay calificaciones
- Papel del gen p53 en la señalización y comunicación celularDocumento5 páginasPapel del gen p53 en la señalización y comunicación celularDaniel AscencioAún no hay calificaciones