También podría gustarte
- Anatomia y Fisiología del Sistema Nervioso IIDe EverandAnatomia y Fisiología del Sistema Nervioso IICalificación: 4 de 5 estrellas4/5 (28)
- Articulo Neuroplasticidad y RehabilitacionDocumento4 páginasArticulo Neuroplasticidad y RehabilitacionPaoliita SangayAún no hay calificaciones
- El nervio periférico: Estructura y funciónDe EverandEl nervio periférico: Estructura y funciónAún no hay calificaciones
- Evaluación PosturalDocumento91 páginasEvaluación PosturalOscar Muñoz Chuquilín100% (1)
- El CerebeloDocumento7 páginasEl Cerebeloquispearapaabraham452Aún no hay calificaciones
- Epilepsia y Cerebelo.Documento14 páginasEpilepsia y Cerebelo.FranciscoAún no hay calificaciones
- EL CerebeloDocumento4 páginasEL CerebelobelenquiuAún no hay calificaciones
- Resumen Primer Parcial Neurofisiologia Iorio 2016Documento35 páginasResumen Primer Parcial Neurofisiologia Iorio 2016Azul AlbertolliAún no hay calificaciones
- TareaDocumento4 páginasTareaYleana RojasAún no hay calificaciones
- Bases Biológicas Del ComportamientoDocumento4 páginasBases Biológicas Del ComportamientoROBIN BRANDON ROJAS ALAVAAún no hay calificaciones
- Bases Biológicas Del ComportamientoDocumento4 páginasBases Biológicas Del ComportamientoROBIN BRANDON ROJAS ALAVAAún no hay calificaciones
- Anatomia Del Sist NerviosoDocumento30 páginasAnatomia Del Sist NerviosoDulce ReyesAún no hay calificaciones
- #4 CerebeloDocumento8 páginas#4 CerebeloMonserrat Olivo TalaveraAún no hay calificaciones
- 4.plasticidad y Aprendizaje PDFDocumento44 páginas4.plasticidad y Aprendizaje PDFAminLopez100% (1)
- Me 2 PDFDocumento22 páginasMe 2 PDFLindsay ParraAún no hay calificaciones
- ¿Qué Es El Cerebelo, Qué Partes Lo Conforman y Qué Función Tiene?Documento11 páginas¿Qué Es El Cerebelo, Qué Partes Lo Conforman y Qué Función Tiene?Beni ColungaAún no hay calificaciones
- Cambiar Tu Cerebro Es PosibleDocumento8 páginasCambiar Tu Cerebro Es PosibleNury CarvajalAún no hay calificaciones
- 8 - MOTOR II - 3.3 - CerebeloDocumento8 páginas8 - MOTOR II - 3.3 - CerebeloFacundo RodriguezAún no hay calificaciones
- El CerebeloDocumento7 páginasEl CerebeloArleen SalinasAún no hay calificaciones
- APUNTE Clase 10Documento12 páginasAPUNTE Clase 10Felipe GuzmanAún no hay calificaciones
- Luria - La Organizacioěn Funcional Del CerebroDocumento21 páginasLuria - La Organizacioěn Funcional Del CerebroNatalia SanchezAún no hay calificaciones
- Reporte de Lectura Neurofisilogía 1Documento3 páginasReporte de Lectura Neurofisilogía 1Héctor Arturo Cruz GranilloAún no hay calificaciones
- El Cerebelo AcvDocumento5 páginasEl Cerebelo Acvdaniel martinezAún no hay calificaciones
- Glosario 2 Angel Figueroa PlascenciaDocumento11 páginasGlosario 2 Angel Figueroa PlascenciaAngel FiguerAún no hay calificaciones
- Centros Reguladores - de La PosturaDocumento2 páginasCentros Reguladores - de La PosturaALEJANDRO OSTOIC ROZZIAún no hay calificaciones
- MesencefaloDocumento17 páginasMesencefaloClinton anderson Luque mamaniAún no hay calificaciones
- Compendio NeurpsicoDocumento26 páginasCompendio NeurpsicodamianAún no hay calificaciones
- Las Bases Biológicas Del Aprendizaje. GlejzerDocumento26 páginasLas Bases Biológicas Del Aprendizaje. GlejzerMarcela RodriguezAún no hay calificaciones
- 23 04 2018 - Guia Fisiología Neuronal Norton Contreras Bases Biológicas y NeurocienciasDocumento27 páginas23 04 2018 - Guia Fisiología Neuronal Norton Contreras Bases Biológicas y NeurocienciasJuan FrancoAún no hay calificaciones
- HipocampoDocumento13 páginasHipocampoKingsley Junior Ibáñez HancoAún no hay calificaciones
- El CerebeloDocumento5 páginasEl CerebeloElena OlivaresAún no hay calificaciones
- Articulo Del Desarrollo Neurologico Del Cerebro y Su Implicancia en El Aprendizaje-1506Documento25 páginasArticulo Del Desarrollo Neurologico Del Cerebro y Su Implicancia en El Aprendizaje-1506A Ma K ReyAún no hay calificaciones
- Apunte 4Documento14 páginasApunte 4Ezequiel MAún no hay calificaciones
- Neuropsicología, Conceptos y TecnicasDocumento26 páginasNeuropsicología, Conceptos y Tecnicasangel esteban palomino bedoyaAún no hay calificaciones
- Neurobiologia Del Movimiento VoluntarioDocumento8 páginasNeurobiologia Del Movimiento VoluntarioAzul MerloAún no hay calificaciones
- Fisio Nervio 1Documento7 páginasFisio Nervio 1Anadella RavalliAún no hay calificaciones
- Estructura Del Cerebro HumanoDocumento12 páginasEstructura Del Cerebro HumanoAlex Fercho Chavez Ladines100% (1)
- Monografia AnatomiaDocumento18 páginasMonografia AnatomiaBeatriz Florian CuevaAún no hay calificaciones
- NeurogenesisDocumento9 páginasNeurogenesisfcovedderAún no hay calificaciones
- Linaje CelularDocumento6 páginasLinaje CelularDiana OrtizAún no hay calificaciones
- Leira 7 PDFDocumento7 páginasLeira 7 PDFPatricia PorrinoAún no hay calificaciones
- Ensayo Del Psicobiogia Sistema NerviosoDocumento8 páginasEnsayo Del Psicobiogia Sistema NerviosoYohana Corzo RodriguezAún no hay calificaciones
- 14 DisejecutivosDocumento36 páginas14 Disejecutivosherminia_ardila50% (2)
- Lectura Complementaria 1-1Documento14 páginasLectura Complementaria 1-1Mily Peña ZuritaAún no hay calificaciones
- Neuroplasticidad O.foster M.terzaghiDocumento16 páginasNeuroplasticidad O.foster M.terzaghinicole50% (2)
- Separata Sesion 2 BasesDocumento12 páginasSeparata Sesion 2 BasesAndré FernándezAún no hay calificaciones
- Me 3Documento13 páginasMe 3Lindsay ParraAún no hay calificaciones
- CEREBELODocumento20 páginasCEREBELOFranny DelgadoAún no hay calificaciones
- Colaborativo 1 - 401504 - 116Documento11 páginasColaborativo 1 - 401504 - 116Jhonatan ToribioAún no hay calificaciones
- Sistema SensitivomotorDocumento3 páginasSistema SensitivomotorMONICA ALCANTARA PAZAún no hay calificaciones
- Aprendizaje y Memoria ImplícitaDocumento12 páginasAprendizaje y Memoria ImplícitaMAR4ITZAMGHAAún no hay calificaciones
- Principios Básicos Del Sistema Nervioso CentralDocumento11 páginasPrincipios Básicos Del Sistema Nervioso CentralYunuelt Velázquez100% (2)
- Untitled (Draft)Documento8 páginasUntitled (Draft)baytianreAndreaAún no hay calificaciones
- MetencéfaloDocumento12 páginasMetencéfaloTatiana Vidal VidalAún no hay calificaciones
- Astrocitoma de Bajo Grado Visualizado Por Resonancia Magnética Cerebral Con Contraste en Paciente Masculino de 24 AnosDocumento14 páginasAstrocitoma de Bajo Grado Visualizado Por Resonancia Magnética Cerebral Con Contraste en Paciente Masculino de 24 AnosFrancisco ChaconAún no hay calificaciones
- Repaso Simulagro de Tres Editores.Documento17 páginasRepaso Simulagro de Tres Editores.Valentina MuñozAún no hay calificaciones
- Anatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2De EverandAnatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2Aún no hay calificaciones
- Clase 1 Estructura CorporalDocumento14 páginasClase 1 Estructura CorporalFrancisco ValenzuelaAún no hay calificaciones
- Células Procariotas y EucariotasDocumento11 páginasCélulas Procariotas y EucariotasRoger DiazAún no hay calificaciones
- Chrisie - Tesis - Titulo - 2017 GuayabaDocumento116 páginasChrisie - Tesis - Titulo - 2017 GuayabaMarcela MolinaAún no hay calificaciones
- Histologia Parcial 2Documento123 páginasHistologia Parcial 2raymery182618Aún no hay calificaciones
- Cuestionario de Lengua y Literatura Del Sexto ParcialDocumento8 páginasCuestionario de Lengua y Literatura Del Sexto ParcialJostyn100% (1)
- Aparato Reproductor Humano para Quinto de SecundariaDocumento5 páginasAparato Reproductor Humano para Quinto de Secundariamibellagenio007Aún no hay calificaciones
- Introduccion A La Fisiologia HumanaDocumento16 páginasIntroduccion A La Fisiologia HumanaevelinmunizAún no hay calificaciones
- IctusDocumento24 páginasIctusELIAún no hay calificaciones
- Momento Optimo de Inseminación Artificial en BovinosDocumento17 páginasMomento Optimo de Inseminación Artificial en BovinosVenezuelaGanadera.comAún no hay calificaciones
- Cuerpos SuperioresDocumento5 páginasCuerpos SuperioresLola PerezAún no hay calificaciones
- DARBEPOETINADocumento10 páginasDARBEPOETINAMateo FigueroaAún no hay calificaciones
- Eje Hipotálamo-Hipófisis-Ovario: o Fase FolicularDocumento5 páginasEje Hipotálamo-Hipófisis-Ovario: o Fase FolicularRosy DíazAún no hay calificaciones
- Cómputos MétricosDocumento3 páginasCómputos MétricosAlfredo DottaviAún no hay calificaciones
- Medio HipotónicoDocumento6 páginasMedio HipotónicoJesus E. Cabezudo Calderon100% (2)
- CONCLUSIONES EDOCRINOLOGíA.7EDDocumento1 páginaCONCLUSIONES EDOCRINOLOGíA.7EDemmAún no hay calificaciones
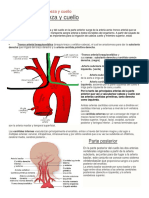
- Irrigación Venosa y Arterial Del Cuello, Cara y CabezaDocumento28 páginasIrrigación Venosa y Arterial Del Cuello, Cara y Cabezasharon100% (1)
- Organos LinfoidesDocumento42 páginasOrganos LinfoidesLicetzita LizAún no hay calificaciones
- ExamenDocumento4 páginasExamenMarcela CorralesAún no hay calificaciones
- La Circulación en Los Seres VivosDocumento10 páginasLa Circulación en Los Seres VivosNoa Natividad100% (1)
- Tangled 4.5 - It's A Wonderful Tangled Christmas CarolDocumento73 páginasTangled 4.5 - It's A Wonderful Tangled Christmas CarolIsrra James AndourAún no hay calificaciones
- Art9 PDFDocumento16 páginasArt9 PDFJenny ReyesAún no hay calificaciones
- Técnicas de Eliminación de Secrecoines PDFDocumento85 páginasTécnicas de Eliminación de Secrecoines PDFBetancourt KarimeAún no hay calificaciones
- Ginecología 1 PDocumento63 páginasGinecología 1 PPau HcAún no hay calificaciones
- Monografía NeurocienciaDocumento6 páginasMonografía NeurocienciaAnibal AlvarezAún no hay calificaciones
- Dominios y ClasesDocumento6 páginasDominios y ClasesJhon Pariacuri MedinaAún no hay calificaciones
- FIcha de TareaDocumento2 páginasFIcha de Tareateacher enderAún no hay calificaciones
- Fisiología NeuronalDocumento16 páginasFisiología NeuronalalfonsoAún no hay calificaciones
- Ibuprofeno Chidaa 1Documento18 páginasIbuprofeno Chidaa 1Dany Chavez50% (2)
- Cuál Es La Importancia de Tener Buenos Hábitos HigiénicosDocumento4 páginasCuál Es La Importancia de Tener Buenos Hábitos HigiénicosPercy E. Távara ArméstarAún no hay calificaciones