También podría gustarte
- Dianas de La Acción FarmacológicaDocumento4 páginasDianas de La Acción FarmacológicaJorge Didier100% (2)
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017De EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Aún no hay calificaciones
- Bioenergética. Introducción a la teoría quimiosmóticaDe EverandBioenergética. Introducción a la teoría quimiosmóticaAún no hay calificaciones
- Certificado LunaDocumento2 páginasCertificado LunaJulian PerezAún no hay calificaciones
- BIOSEÑALIZACIÓNDocumento11 páginasBIOSEÑALIZACIÓNelisa ponce de leonAún no hay calificaciones
- Monografia Equipo6 FarmacodinamiaDocumento2 páginasMonografia Equipo6 FarmacodinamiaTerra NewAún no hay calificaciones
- Unicación IntracelularDocumento5 páginasUnicación IntracelularAzura YouhichiAún no hay calificaciones
- Videoconferencia FarmacodinamiaDocumento16 páginasVideoconferencia FarmacodinamiaIsaAún no hay calificaciones
- Señalizacion Celular 2 PDFDocumento56 páginasSeñalizacion Celular 2 PDFElva EstefaneroAún no hay calificaciones
- Catalisis y Reaccion de AcoplamientoDocumento7 páginasCatalisis y Reaccion de AcoplamientoMarley CajamarcaAún no hay calificaciones
- Seminario Receptores FisiologíaDocumento12 páginasSeminario Receptores FisiologíaGilmar Daniel MincholaAún no hay calificaciones
- GlucólisisDocumento7 páginasGlucólisisDaniel EduardoAún no hay calificaciones
- 6 - Teoría - Vereau - ReceptoresDocumento11 páginas6 - Teoría - Vereau - ReceptoresSol CamusAún no hay calificaciones
- Tipos de Receptores Celulares y Mecanismos de TransducciónDocumento38 páginasTipos de Receptores Celulares y Mecanismos de TransducciónKariem Noelia Sherezade Siquero Vera100% (1)
- Unidad 1 - Clase 2 - Membrana - Transporte y Transducción de SeñalesDocumento27 páginasUnidad 1 - Clase 2 - Membrana - Transporte y Transducción de SeñalesAntonia Samur ContrerasAún no hay calificaciones
- S03C05 - Membrana (III)Documento42 páginasS03C05 - Membrana (III)Alejandro B. SilvaAún no hay calificaciones
- Biología - Módulo 02 - Señalización y Comunicación Celular PDFDocumento22 páginasBiología - Módulo 02 - Señalización y Comunicación Celular PDFGeraldine CastiñeiraAún no hay calificaciones
- Apuntes de FarmaDocumento6 páginasApuntes de FarmaAndrea AlriviAún no hay calificaciones
- Bioquimica - Receptores de MembranaDocumento6 páginasBioquimica - Receptores de MembranaGeison MartinsAún no hay calificaciones
- Resumen Capitulo 4Documento5 páginasResumen Capitulo 4Magally Castañeda GómezAún no hay calificaciones
- Señalización CelularDocumento13 páginasSeñalización CelularAmadeo CabelloAún no hay calificaciones
- Temas 10 Al 14 ResumenDocumento19 páginasTemas 10 Al 14 ResumenClaudia MeléndezAún no hay calificaciones
- 02-FARMACODINAMIA ResumenDocumento11 páginas02-FARMACODINAMIA ResumenMaxy RosalesAún no hay calificaciones
- Comunicación CelularDocumento29 páginasComunicación CelularMauricio OchoaAún no hay calificaciones
- 2 - Transducción de SeñalesDocumento5 páginas2 - Transducción de SeñalesHernán GuietAún no hay calificaciones
- 6 Metabolismo Celular. Recepción y Traducción de SeñalesDocumento10 páginas6 Metabolismo Celular. Recepción y Traducción de Señaleslizbeth TibanlomboAún no hay calificaciones
- Señalizacion CelularDocumento4 páginasSeñalizacion CelularVanesa Soledad RomanoAún no hay calificaciones
- Prueba 3 BioquímicaDocumento52 páginasPrueba 3 BioquímicaKimmiiNadalesAún no hay calificaciones
- C2 Farmacodinamia 11-08-22Documento6 páginasC2 Farmacodinamia 11-08-22Sebastian Ignacio Isla PastenAún no hay calificaciones
- Farmacologia MolecularDocumento5 páginasFarmacologia MolecularolgakatherineAún no hay calificaciones
- PBDocumento7 páginasPBLUCIANA ABIGAIL TORRES MUCHAAún no hay calificaciones
- Taller de Repaso Bioseñalización y Bioenergética Marzo 2020Documento13 páginasTaller de Repaso Bioseñalización y Bioenergética Marzo 2020mafe ospinaAún no hay calificaciones
- Resumen Neuro 2Documento24 páginasResumen Neuro 2Constanza BoveraAún no hay calificaciones
- Transducción de Señales - Fisiología IDocumento21 páginasTransducción de Señales - Fisiología IKeka Quijano100% (1)
- Señalizacion CelularDocumento12 páginasSeñalizacion CelularYean Paul Cruz SuarezAún no hay calificaciones
- Mapa Conceptual Mecanismos Señalizacion TransmembranalDocumento1 páginaMapa Conceptual Mecanismos Señalizacion Transmembranalale steph33% (3)
- Tipos de Receptores, Pregunta 2 de FarmacodinamiaDocumento3 páginasTipos de Receptores, Pregunta 2 de FarmacodinamiaDaniela HernandezAún no hay calificaciones
- Guia de SinapsisDocumento5 páginasGuia de SinapsisCarolina Pastén AlvarezAún no hay calificaciones
- Clasificacion de Receptores Segun Mecanismo de AccionDocumento6 páginasClasificacion de Receptores Segun Mecanismo de AccionDeinir SolanoAún no hay calificaciones
- Farmacología MolecularDocumento35 páginasFarmacología MolecularXIMENA LISSET HERNANDEZ VALDEZAún no hay calificaciones
- Receptores de MembranaDocumento28 páginasReceptores de MembranaAngela AliagaAún no hay calificaciones
- Senalizacion IntercelularDocumento35 páginasSenalizacion IntercelularIrving ParraAún no hay calificaciones
- 4 Receptores de MembranaDocumento32 páginas4 Receptores de MembranaDhara RicciAún no hay calificaciones
- FarmacodinamiaDocumento59 páginasFarmacodinamiaRodrigo S. AvilésAún no hay calificaciones
- Mecanismos Generales de Comunicación CelularDocumento4 páginasMecanismos Generales de Comunicación CelularPatricia SolanoAún no hay calificaciones
- Señalización Celular, Camilo Escobedo Fernández PDFDocumento5 páginasSeñalización Celular, Camilo Escobedo Fernández PDFcamiloesferAún no hay calificaciones
- UNIDAD IX. Señalización CelularDocumento105 páginasUNIDAD IX. Señalización CelularHelena QuintanaAún no hay calificaciones
- Transducción de Señales Flopis PDFDocumento19 páginasTransducción de Señales Flopis PDFFlorencia PeñaAún no hay calificaciones
- Transducción de SeñalesDocumento11 páginasTransducción de SeñalesMartina Goñi PadrónAún no hay calificaciones
- DINAMICA 4 (Apuntes)Documento2 páginasDINAMICA 4 (Apuntes)Juan FernándezAún no hay calificaciones
- Enzimas FarmaDocumento19 páginasEnzimas FarmaAlejandra VegaAún no hay calificaciones
- Mecanismo Generales de Señalización Celular - 240411 - 110457Documento3 páginasMecanismo Generales de Señalización Celular - 240411 - 110457Jose ArayaAún no hay calificaciones
- Receptores FarmacológicosDocumento13 páginasReceptores FarmacológicosCinthia Diaz100% (1)
- 02 - Funcion de Proteinas 2017 PDFDocumento150 páginas02 - Funcion de Proteinas 2017 PDFAnthony VargasAún no hay calificaciones
- Tema 8-Intro Met AlumnosDocumento28 páginasTema 8-Intro Met AlumnosJesús María Martínez PinillaAún no hay calificaciones
- Act 6Documento1 páginaAct 6Karina Berry MendozaAún no hay calificaciones
- Fisiologia Semana 4Documento11 páginasFisiologia Semana 4nicollgarsomAún no hay calificaciones
- Resumen HORMONASDocumento8 páginasResumen HORMONASmartinamorelvulliez12345Aún no hay calificaciones
- Videos Farmacodinamia Completo (Avirtual)Documento17 páginasVideos Farmacodinamia Completo (Avirtual)Melany GallardoAún no hay calificaciones
- Tema 2 (Parte 2)Documento6 páginasTema 2 (Parte 2)daliasandulacheAún no hay calificaciones
- Los Seres Vivos Dependen De La Mecánica CuánticaDe EverandLos Seres Vivos Dependen De La Mecánica CuánticaAún no hay calificaciones
- Unidad 4. Metabolismo Lipidos 2Documento18 páginasUnidad 4. Metabolismo Lipidos 2Jessica RodríguezAún no hay calificaciones
- Unidad 4. Metabolismo Lipidos 1Documento16 páginasUnidad 4. Metabolismo Lipidos 1Jessica RodríguezAún no hay calificaciones
- Unidad 2. Metabolismo de Carbohidratos 3Documento7 páginasUnidad 2. Metabolismo de Carbohidratos 3Jessica RodríguezAún no hay calificaciones
- Unidad 2. Metabolismo de Carbohidratos 1Documento12 páginasUnidad 2. Metabolismo de Carbohidratos 1Jessica RodríguezAún no hay calificaciones
- Solvente Dieléctrico SS-25 NCDocumento5 páginasSolvente Dieléctrico SS-25 NCGabriel Flores33% (3)
- Proyecto Carter Hileradora FinalDocumento57 páginasProyecto Carter Hileradora FinalMj BmAún no hay calificaciones
- Info07 Actuación Situación Riesgo EscuelaDocumento1 páginaInfo07 Actuación Situación Riesgo EscuelaRodrigo Rebollar León100% (2)
- Documento Calculos de Juego MecanicoDocumento33 páginasDocumento Calculos de Juego MecanicoMarvin NajarroAún no hay calificaciones
- Dificultad, Trastorno, DiscapacidadDocumento2 páginasDificultad, Trastorno, DiscapacidadJMancilla9Aún no hay calificaciones
- Nota para Políticas Sociales 3Documento20 páginasNota para Políticas Sociales 3Secretaría de la Integración Social CentroamericanaAún no hay calificaciones
- Infografia Sem 3Documento1 páginaInfografia Sem 3A. GrisAún no hay calificaciones
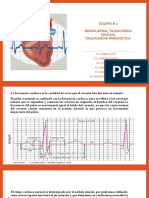
- Bradicardia Taquicardia Equipo #2Documento11 páginasBradicardia Taquicardia Equipo #2Alberto VazquezAún no hay calificaciones
- Biogas A Partir de PollinazaDocumento15 páginasBiogas A Partir de Pollinazaledatintaya100% (1)
- Uso de Repelente NaturalDocumento11 páginasUso de Repelente NaturalGermán DÍAZAún no hay calificaciones
- Administracion Por Via Im-Sc-IdDocumento31 páginasAdministracion Por Via Im-Sc-IdLaura Daniela Tovar CamachoAún no hay calificaciones
- Celula 01Documento3 páginasCelula 01Saryss notessAún no hay calificaciones
- Intubación EndotraquealDocumento17 páginasIntubación Endotraquealmaria100% (1)
- Actividad Física y SaludDocumento21 páginasActividad Física y SaludGisbellAcostaVillasmilAún no hay calificaciones
- Identificacion de PCCDocumento5 páginasIdentificacion de PCCRuth Sofia Aguilar CcolqqueAún no hay calificaciones
- Actividad 1 CarnesDocumento4 páginasActividad 1 CarnesTatihana Blanco RinconAún no hay calificaciones
- La Investigación en PsicologíaDocumento52 páginasLa Investigación en PsicologíajesshernAún no hay calificaciones
- Preguntas de HidrologíaDocumento3 páginasPreguntas de HidrologíaEtzel Ed MejAún no hay calificaciones
- Examen Casero 2Documento6 páginasExamen Casero 2Nora NuñezAún no hay calificaciones
- Cuadro de VitaminasDocumento6 páginasCuadro de VitaminasAndrea Parada100% (1)
- Liquidacion Servicios Publicos IquemDocumento2 páginasLiquidacion Servicios Publicos IquemRosa MejiaAún no hay calificaciones
- RAP 135 Capitulo C - Instrumentos y EquiposDocumento17 páginasRAP 135 Capitulo C - Instrumentos y Equipossergiolopez23Aún no hay calificaciones
- AlgasDocumento3 páginasAlgasNicolle Andrea Morales CárdenasAún no hay calificaciones
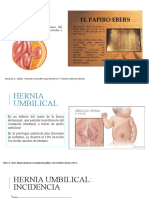
- HerniasDocumento44 páginasHerniasJORGE DIAZAún no hay calificaciones
- Carbopol y Excipiente Acofar Gel Carb MeroDocumento4 páginasCarbopol y Excipiente Acofar Gel Carb MeroDavidMiñanPacheco0% (1)
- Matriz de Impacto CruzadoDocumento6 páginasMatriz de Impacto CruzadoStephanie RojasAún no hay calificaciones
- Tutoría 2 HábitosDocumento3 páginasTutoría 2 HábitosAndrey PCAún no hay calificaciones
- Veronica Mata Alvarez - 5AFPDocumento24 páginasVeronica Mata Alvarez - 5AFPVer019Aún no hay calificaciones
- Normatividad en Salud OcupacionalDocumento38 páginasNormatividad en Salud OcupacionalleoAún no hay calificaciones