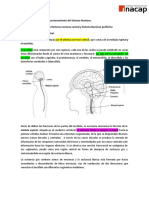
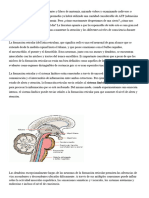
También podría gustarte
- 20 Reglas de BasquetbolDocumento2 páginas20 Reglas de BasquetbolRosyLeyva78% (65)
- Sistema NerviosoDocumento2 páginasSistema NerviosoK Susana M Bazan100% (1)
- Anatomía y Fisiología del Sistema NerviosoDe EverandAnatomía y Fisiología del Sistema NerviosoCalificación: 4 de 5 estrellas4/5 (19)
- Vida SaludableDocumento3 páginasVida SaludableRosyLeyvaAún no hay calificaciones
- ANATOMOFISILOGÍA II - Resumen para El FinalDocumento85 páginasANATOMOFISILOGÍA II - Resumen para El Finalcandela rodriguez gherbazAún no hay calificaciones
- Diviana Rocca Seccion B Neurofisiologia y Fisiopatologia Unidad 3Documento114 páginasDiviana Rocca Seccion B Neurofisiologia y Fisiopatologia Unidad 3oriana roccaAún no hay calificaciones
- SIstema Nervioso Periferico (SNP)Documento12 páginasSIstema Nervioso Periferico (SNP)Iliana DiamondAún no hay calificaciones
- NeurotransmisiónDocumento8 páginasNeurotransmisiónvtaglemendoza5223Aún no hay calificaciones
- Sistema Nervioso SimpaticoDocumento20 páginasSistema Nervioso SimpaticoLina VelezAún no hay calificaciones
- CDocumento7 páginasCLuis AlejosAún no hay calificaciones
- Resumen de Generalidades Del Sistema Nervioso.Documento7 páginasResumen de Generalidades Del Sistema Nervioso.Adriana ClarosAún no hay calificaciones
- Funciones Del Sistema Nervioso Central y PeriféricoDocumento6 páginasFunciones Del Sistema Nervioso Central y PerifériconadiaAún no hay calificaciones
- Sist AutonomoDocumento16 páginasSist AutonomoSofíaAún no hay calificaciones
- SNA - ApunteDocumento8 páginasSNA - ApuntemariaAún no hay calificaciones
- Leccion 10Documento5 páginasLeccion 10pm2rn4vdm8Aún no hay calificaciones
- Biologia Del Movimiento II (Sist. Nervioso)Documento17 páginasBiologia Del Movimiento II (Sist. Nervioso)Joaquin GorositoAún no hay calificaciones
- Ao 10-2Documento10 páginasAo 10-2Una patata Llamada AntonellaAún no hay calificaciones
- Tarea Nro 2 Ao 2 MFH II Semana 2 5 Al 10 de Julio de 2021Documento4 páginasTarea Nro 2 Ao 2 MFH II Semana 2 5 Al 10 de Julio de 2021Prof. Jose MurilloAún no hay calificaciones
- Tema 1. Sistema Nervioso AutónomoDocumento11 páginasTema 1. Sistema Nervioso AutónomoEudimarAún no hay calificaciones
- Neuroanatomía. Dr. Andrés González GarridoDocumento26 páginasNeuroanatomía. Dr. Andrés González Garridojose jaime botello valle100% (1)
- Aspectos Generales Del Sistema NerviosoDocumento6 páginasAspectos Generales Del Sistema NerviosoJuan CuevaAún no hay calificaciones
- MonografiaDocumento14 páginasMonografiaFrancisco Lodigiani100% (1)
- Sistema NerviosoDocumento12 páginasSistema NerviosoGabriela Sofia PacAún no hay calificaciones
- Sistema NerviosoDocumento10 páginasSistema NerviosoNicoAún no hay calificaciones
- Informe Anatomia IiDocumento7 páginasInforme Anatomia IijhosmairimrodriguezAún no hay calificaciones
- Trabajo Sistema NerviosoDocumento15 páginasTrabajo Sistema NerviosoIvonne PontonAún no hay calificaciones
- Sistema Nervioso Autónomo IDocumento5 páginasSistema Nervioso Autónomo ILRI19040 - CECILIA GARCIA NI�OAún no hay calificaciones
- Anat Aplic 1º Bach T2 Coord NervDocumento24 páginasAnat Aplic 1º Bach T2 Coord Nervjuancarloscamposromero173Aún no hay calificaciones
- Generalidades Anatómicas y FisiológicasDocumento60 páginasGeneralidades Anatómicas y Fisiológicaspaula740003Aún no hay calificaciones
- Formacion ReticularDocumento11 páginasFormacion Reticularvargassahori30Aún no hay calificaciones
- Ficha TP6 Sistema Nervioso IIDocumento16 páginasFicha TP6 Sistema Nervioso IIJesús GiménezAún no hay calificaciones
- Presentación1 Sistema NerviosoDocumento27 páginasPresentación1 Sistema NerviosoJose Guillermo Herrejon DelgadilloAún no hay calificaciones
- Organografía Microscópica HumanaDocumento144 páginasOrganografía Microscópica HumanaCristóbal Muñoz Pérez100% (2)
- Trabajo Final de AnatomiaDocumento15 páginasTrabajo Final de AnatomiaJose Luis Hernandez100% (1)
- M1 T6 - NeuroPsDocumento13 páginasM1 T6 - NeuroPsThonny EspinosaAún no hay calificaciones
- Sistema Nervioso E.Y.S.LDocumento23 páginasSistema Nervioso E.Y.S.LMarco Antonio Urias SalgadoAún no hay calificaciones
- Comunicacion Neuroral Tema2Documento6 páginasComunicacion Neuroral Tema2Miguel Antonio NMMAún no hay calificaciones
- Anatomía Del Sistema NerviosoDocumento31 páginasAnatomía Del Sistema NerviosoBianka NavarreteAún no hay calificaciones
- Modulo Neuroanatomía Ufro 2 PDFDocumento230 páginasModulo Neuroanatomía Ufro 2 PDFcristian moragaAún no hay calificaciones
- ANTONIO NIÑO CRISTIAN ALEJANDRO. GRUPO3 AF Trabajo 2 Fisio AnimDocumento9 páginasANTONIO NIÑO CRISTIAN ALEJANDRO. GRUPO3 AF Trabajo 2 Fisio AnimCristian AntonioAún no hay calificaciones
- Sistema NerviosoDocumento12 páginasSistema NerviosoAngel AponteAún no hay calificaciones
- Cordon Neural - Fisiología AnimalDocumento13 páginasCordon Neural - Fisiología AnimalSein SaggAún no hay calificaciones
- P 9 Szmmdfsda &expires 1215573344&awsaccesskeyid 1 Z 5 T 9 H 8 PQ 39 V 6 F 79 V 8 G 2Documento56 páginasP 9 Szmmdfsda &expires 1215573344&awsaccesskeyid 1 Z 5 T 9 H 8 PQ 39 V 6 F 79 V 8 G 2api-3735753Aún no hay calificaciones
- Apuntes NeuroanatomíaDocumento109 páginasApuntes NeuroanatomíaMichelle Acuña EspinozaAún no hay calificaciones
- Monografia Sistema NerviosoDocumento13 páginasMonografia Sistema NerviosoCiber netAún no hay calificaciones
- Vegetativo 2013Documento18 páginasVegetativo 2013Amaya ArnaizAún no hay calificaciones
- Anatomia y Fisiologia Sistema NerviosoDocumento10 páginasAnatomia y Fisiologia Sistema NerviosoCamila Alvarez MayorgaAún no hay calificaciones
- CURTIS H. - Capítulo 31 - U3Documento6 páginasCURTIS H. - Capítulo 31 - U3Iaia RuizAún no hay calificaciones
- Sistema Nervioso Entérico y La Motilidad GastrointestinalDocumento5 páginasSistema Nervioso Entérico y La Motilidad GastrointestinalMileydy AlvaradoAún no hay calificaciones
- .E.A. Guido VillagomezDocumento5 páginas.E.A. Guido VillagomezMarcelo ChavezAún no hay calificaciones
- Introduccion Basica VV 0.202110 PDFDocumento19 páginasIntroduccion Basica VV 0.202110 PDFJeannette del RosarioAún no hay calificaciones
- GeneralidadessistnervDocumento22 páginasGeneralidadessistnervJuan OjedaAún no hay calificaciones
- Desarrollo Del Sistema Nervioso Periferico 4ta Unidad TerminadoDocumento20 páginasDesarrollo Del Sistema Nervioso Periferico 4ta Unidad TerminadoAlexandra Yacely Perez GamonalAún no hay calificaciones
- (Haines) Introducción A La Estructura Del Sistema Nervioso CentralDocumento5 páginas(Haines) Introducción A La Estructura Del Sistema Nervioso CentralCesar EgusquizaAún no hay calificaciones
- Procesos Biológicos I, Guía 2Documento30 páginasProcesos Biológicos I, Guía 2Angela KKAún no hay calificaciones
- Sistema NerviosoDocumento70 páginasSistema NerviosoMaJo Montoya100% (1)
- Neurociencias II (SUMATIVA 3) Diana RivasDocumento10 páginasNeurociencias II (SUMATIVA 3) Diana RivasDiana RivasAún no hay calificaciones
- Valoración y Cuidados Enfermeros Al Paciente Con Problemas NeurológicosDocumento160 páginasValoración y Cuidados Enfermeros Al Paciente Con Problemas NeurológicoswallatoallaAún no hay calificaciones
- Sistema Nervioso y Endocrino PDFDocumento28 páginasSistema Nervioso y Endocrino PDFGloria Sarmiento Fuertes75% (4)
- Leccion 1Documento8 páginasLeccion 1pm2rn4vdm8Aún no hay calificaciones
- Anatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2De EverandAnatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2Aún no hay calificaciones
- El cannabis en patologías del sistema nervioso centralDe EverandEl cannabis en patologías del sistema nervioso centralAún no hay calificaciones
- Mate1 REFORZAMIENTO ALUMNODocumento13 páginasMate1 REFORZAMIENTO ALUMNORosyLeyva100% (1)
- Infografia de Lengua MaternaDocumento1 páginaInfografia de Lengua MaternaRosyLeyvaAún no hay calificaciones
- Alimentos y Su Aporte CalóricoDocumento2 páginasAlimentos y Su Aporte CalóricoRosyLeyvaAún no hay calificaciones
- Origen y Evolución Del InternetDocumento5 páginasOrigen y Evolución Del InternetRosyLeyvaAún no hay calificaciones
- Consigna 2: Consideren La Situación de La Consigna 1, Con La Diferencia de Que El Lado Correspondiente A 9Documento7 páginasConsigna 2: Consideren La Situación de La Consigna 1, Con La Diferencia de Que El Lado Correspondiente A 9RosyLeyva0% (1)
- Clase MaquinasDocumento14 páginasClase MaquinasRosyLeyvaAún no hay calificaciones
- Delegar IDocumento3 páginasDelegar IRosyLeyvaAún no hay calificaciones
- 02 Bimestreapuntesdeinformatica 1 CienciaytecnologiaDocumento10 páginas02 Bimestreapuntesdeinformatica 1 CienciaytecnologiaRosyLeyvaAún no hay calificaciones
- Proceso de Técnico Del Papel BondDocumento6 páginasProceso de Técnico Del Papel BondRosyLeyva100% (1)
- Surgimiento Del SindicalismoDocumento72 páginasSurgimiento Del SindicalismoRosyLeyva100% (1)
- Delegacion de FuncionesDocumento1 páginaDelegacion de FuncionesRosyLeyvaAún no hay calificaciones
- Maquina CompuestaDocumento2 páginasMaquina CompuestaRosyLeyvaAún no hay calificaciones
- Diferencias Entre Maquinas Simples y CompuestasDocumento2 páginasDiferencias Entre Maquinas Simples y CompuestasRosyLeyva80% (5)
- Especies EndémicasDocumento4 páginasEspecies EndémicasRosyLeyvaAún no hay calificaciones
- Funciones de Las HerramientasDocumento2 páginasFunciones de Las HerramientasRosyLeyvaAún no hay calificaciones
- Adaptación de La Teoría de Dorothea Orem A Personas ConDocumento13 páginasAdaptación de La Teoría de Dorothea Orem A Personas ConIris Jaquelin RendonAún no hay calificaciones
- 348 Fichas EDITABLE PDFDocumento2 páginas348 Fichas EDITABLE PDFfernando830609Aún no hay calificaciones
- Cargos Ofrecidos Concurso Residencias 2024Documento3 páginasCargos Ofrecidos Concurso Residencias 2024Daniel AragonAún no hay calificaciones
- Casos Clinicos en Imagenes CTODocumento151 páginasCasos Clinicos en Imagenes CTOFreddy Chacón100% (8)
- Cuadro Comparativo Enfoques PsicologicosDocumento7 páginasCuadro Comparativo Enfoques Psicologicosjorge luis bran bedoyaAún no hay calificaciones
- EXOFTALMIADocumento2 páginasEXOFTALMIAAndres ChachaAún no hay calificaciones
- Guardias Ad - Rotación 100721Documento12 páginasGuardias Ad - Rotación 100721emilio moyaoAún no hay calificaciones
- Formato Acciones de MejoraDocumento2 páginasFormato Acciones de MejoraJuanex VzAún no hay calificaciones
- Algoritmo TEPDocumento2 páginasAlgoritmo TEPMICHAEL STICK NARVAEZ CARDENASAún no hay calificaciones
- 1.POE INGRESO DE PERSONAS, OBJETOS Y VEHÍCULOS (07 - 10 - 14) COPIAAAdocxDocumento4 páginas1.POE INGRESO DE PERSONAS, OBJETOS Y VEHÍCULOS (07 - 10 - 14) COPIAAAdocxAndres CamargoAún no hay calificaciones
- Clase 1Documento22 páginasClase 1Rosita MonzonAún no hay calificaciones
- Actividades y Tareas de Alto RiesgoDocumento1 páginaActividades y Tareas de Alto RiesgoJonathan Manjarres100% (1)
- Entrevista Personal IhssDocumento3 páginasEntrevista Personal IhssOskarr Valle100% (1)
- Historia, Efectos de La TMODocumento46 páginasHistoria, Efectos de La TMOAngel López100% (2)
- GeneralidadesDocumento11 páginasGeneralidadesDelia TicllasucaAún no hay calificaciones
- Test de Relaciones Objetales o TRO de PhillipsonDocumento12 páginasTest de Relaciones Objetales o TRO de PhillipsontulayevaiAún no hay calificaciones
- Analisis Proximal y Contenido de Hierro y Calcio de Nostoc Sphaericum "Cushuro" Deshidratado Procedente de La Laguna de Conococha, Catac - HuarazDocumento13 páginasAnalisis Proximal y Contenido de Hierro y Calcio de Nostoc Sphaericum "Cushuro" Deshidratado Procedente de La Laguna de Conococha, Catac - Huarazana benaventeAún no hay calificaciones
- Cedula Sospecha Cáncer en - 18 Años 1er Niv 2016Documento3 páginasCedula Sospecha Cáncer en - 18 Años 1er Niv 2016Alexis PatAún no hay calificaciones
- Centro Hogar GeriatricoDocumento155 páginasCentro Hogar GeriatricoEmily CevallosAún no hay calificaciones
- Mapa Conceptual OsteoporosisDocumento1 páginaMapa Conceptual OsteoporosisJHONAR ALEX OSWALDO VASQUEZ LLANOSAún no hay calificaciones
- Infografia Arroz FortificadoDocumento1 páginaInfografia Arroz FortificadoAmareto123Aún no hay calificaciones
- RotuladoDocumento27 páginasRotuladojuuvazquez040627Aún no hay calificaciones
- 049 2111 060813t01 16000 PDFDocumento767 páginas049 2111 060813t01 16000 PDFGab BacciAún no hay calificaciones
- Normas de Convivencia 22 ActualDocumento3 páginasNormas de Convivencia 22 Actualhermila ruizAún no hay calificaciones
- Trabajo Aptitud FisicaDocumento12 páginasTrabajo Aptitud FisicaKenny Yuliana SeijasAún no hay calificaciones
- Eutanasia Distanasia y OrtotanasiaDocumento22 páginasEutanasia Distanasia y OrtotanasiaOskar Vargas100% (1)
- Diagnóstico Annie 1Documento5 páginasDiagnóstico Annie 1Jorge Armando Ustoa SigalaAún no hay calificaciones
- CINEANTROPOMETRÍADocumento14 páginasCINEANTROPOMETRÍACamilo MoralesAún no hay calificaciones
- Lectura-Línea de InvestigaciónDocumento3 páginasLectura-Línea de Investigaciónluis alberto huillcas enriquezAún no hay calificaciones
- Productos Con Tarifa ReducidaDocumento427 páginasProductos Con Tarifa ReducidaEmmanuel MejiasAún no hay calificaciones