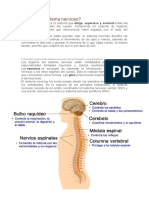
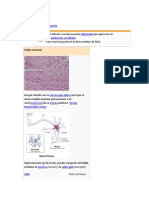
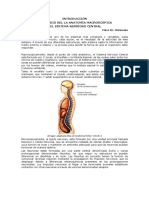
También podría gustarte
- Ensayo de Sistema Nervioso Central y Periferico ActualizadoDocumento8 páginasEnsayo de Sistema Nervioso Central y Periferico ActualizadoAdrian Rivera40% (5)
- Monografia, Sistema NerviosoDocumento22 páginasMonografia, Sistema NerviosoDiego Alejandro Lara Q78% (9)
- Presentación Taller Modulo 1 AutoconocimientoDocumento36 páginasPresentación Taller Modulo 1 AutoconocimientoLuis RamirezAún no hay calificaciones
- Servicio de Ventilacion Mina Huaron PDFDocumento118 páginasServicio de Ventilacion Mina Huaron PDFAlber LoayzaAún no hay calificaciones
- Prac. DIRI (3) PDFDocumento15 páginasPrac. DIRI (3) PDFIsmael Escobar AlvaAún no hay calificaciones
- Sistema NerviosoDocumento10 páginasSistema NerviosoNicoAún no hay calificaciones
- Bases Neuroficiologicas Del AprendizajeDocumento6 páginasBases Neuroficiologicas Del AprendizajeNatalia FariñaAún no hay calificaciones
- Sistema NerviosoDocumento11 páginasSistema Nerviososaulcarvajal23Aún no hay calificaciones
- Biologia Del Movimiento II (Sist. Nervioso)Documento17 páginasBiologia Del Movimiento II (Sist. Nervioso)Joaquin GorositoAún no hay calificaciones
- Tema 8 ANATOMOFISIOLOGIA Y PATOLOGÍA DEL SISTEMA NERVIOSO Y LOS ÓRGANOS DE LOS SENTIDOSDocumento23 páginasTema 8 ANATOMOFISIOLOGIA Y PATOLOGÍA DEL SISTEMA NERVIOSO Y LOS ÓRGANOS DE LOS SENTIDOSalejandraAún no hay calificaciones
- Guia 4 ListaDocumento13 páginasGuia 4 ListaYazmin loraAún no hay calificaciones
- 789unidad 8Documento53 páginas789unidad 8Maria González FariñasAún no hay calificaciones
- Sistema NerviosoDocumento14 páginasSistema NerviosoAlexander GomezAún no hay calificaciones
- Sistema NerviosoDocumento5 páginasSistema NerviosoKARENAún no hay calificaciones
- Sistema Nervioso Contenido, 2023Documento10 páginasSistema Nervioso Contenido, 2023Monica RodriguezAún no hay calificaciones
- Sistema NerviosoDocumento45 páginasSistema NerviosoFrancisco Montecinos100% (1)
- Unidad 13 - M3 - CITEDocumento33 páginasUnidad 13 - M3 - CITEjuanAún no hay calificaciones
- Tema 5 SNDocumento13 páginasTema 5 SNBethaniaAún no hay calificaciones
- Sistema Nervioso IIDocumento10 páginasSistema Nervioso IIItzel Altamirano HernándezAún no hay calificaciones
- Tarea 2 Psicologia GeneralDocumento7 páginasTarea 2 Psicologia GeneralMiguel MontillaAún no hay calificaciones
- MonografiaDocumento14 páginasMonografiaFrancisco Lodigiani100% (1)
- Sistema Nervioso. GeneralidadesDocumento59 páginasSistema Nervioso. Generalidadesgwbn6qhgt9Aún no hay calificaciones
- Desarrollo Del Sistema Nervioso Periferico 4ta Unidad TerminadoDocumento20 páginasDesarrollo Del Sistema Nervioso Periferico 4ta Unidad TerminadoAlexandra Yacely Perez GamonalAún no hay calificaciones
- Sistema Nervioso PerifericoDocumento14 páginasSistema Nervioso PerifericoCarlos Arana MendezAún no hay calificaciones
- Sistema NerviosoDocumento12 páginasSistema NerviosoAngel AponteAún no hay calificaciones
- Sistema Nervioso 2 DiciembreDocumento7 páginasSistema Nervioso 2 DiciembreNathalie Fernanda ʚïɞAún no hay calificaciones
- Qué Es El Sistema NerviosoDocumento6 páginasQué Es El Sistema NerviosoElba100% (1)
- Modulo Neuroanatomía Ufro 2 PDFDocumento230 páginasModulo Neuroanatomía Ufro 2 PDFcristian moragaAún no hay calificaciones
- Fisiologia General y Del Ejercicio Act 1Documento7 páginasFisiologia General y Del Ejercicio Act 1AlissonAún no hay calificaciones
- Presentación1 Sistema NerviosoDocumento27 páginasPresentación1 Sistema NerviosoJose Guillermo Herrejon DelgadilloAún no hay calificaciones
- Tema en ProcesoDocumento11 páginasTema en ProcesoPatricia YZAún no hay calificaciones
- Utel Sistemanervioso S1Documento8 páginasUtel Sistemanervioso S1josselynguizado194Aún no hay calificaciones
- Anat Aplic 1º Bach T2 Coord NervDocumento24 páginasAnat Aplic 1º Bach T2 Coord Nervjuancarloscamposromero173Aún no hay calificaciones
- Trabajo PracticoDocumento8 páginasTrabajo PracticoVanina NavarreteAún no hay calificaciones
- Af1-Mod 5-NervDocumento34 páginasAf1-Mod 5-NervGustavo CamaraAún no hay calificaciones
- Sistema Nervioso E.Y.S.LDocumento23 páginasSistema Nervioso E.Y.S.LMarco Antonio Urias SalgadoAún no hay calificaciones
- Resumen 2do ParcialDocumento18 páginasResumen 2do ParcialmalAún no hay calificaciones
- Sistema Nerviosos 1Documento20 páginasSistema Nerviosos 1esmeraldaAún no hay calificaciones
- Sistema NerviosoDocumento8 páginasSistema NerviosoJuliana AcevedoAún no hay calificaciones
- Tema 1organización Del Sistema Nervioso CentralDocumento32 páginasTema 1organización Del Sistema Nervioso Centralroxana celisAún no hay calificaciones
- Resumen Completo Angel Taveras NeuroanatomiaDocumento45 páginasResumen Completo Angel Taveras NeuroanatomiaElupine PerezAún no hay calificaciones
- MORFOLOGIA Y FUNCION II Sistema NerviosoDocumento56 páginasMORFOLOGIA Y FUNCION II Sistema NerviosoDaniela Garcés QuirogaAún no hay calificaciones
- Parte 2 S.NDocumento36 páginasParte 2 S.NalbertoAún no hay calificaciones
- Exposicion de FisiologiaDocumento5 páginasExposicion de FisiologiaCarmenAún no hay calificaciones
- Sistema NerviosoDocumento5 páginasSistema Nerviosoputo el que lo leaAún no hay calificaciones
- Guía Grado 9 Sistema NerviosoDocumento5 páginasGuía Grado 9 Sistema Nerviosogustavo herreraAún no hay calificaciones
- 6 Sistema NerviosoDocumento54 páginas6 Sistema NerviosoXavi Moran100% (1)
- Sistema NerviosoDocumento26 páginasSistema NerviosoRaquel Correa Chavarria100% (2)
- Ciencias 6Documento4 páginasCiencias 6Patricio RiquelmeAún no hay calificaciones
- El Sistema Nervioso y EndocrinoDocumento6 páginasEl Sistema Nervioso y EndocrinoPedroPerezPerezAún no hay calificaciones
- Tejido NerviosoDocumento18 páginasTejido NerviosoLuzmaria Escalona VargasAún no hay calificaciones
- Sistema NerviosoDocumento20 páginasSistema NerviosoAndrea L. CernaAún no hay calificaciones
- Sistema NerviosoDocumento12 páginasSistema Nerviosoantonella cibotariAún no hay calificaciones
- Introduccion Basica VV 0.202110 PDFDocumento19 páginasIntroduccion Basica VV 0.202110 PDFJeannette del RosarioAún no hay calificaciones
- Texto 1 CARACTERISTICAS - BASICAS - DEL - SISTEMA - NERVIOSODocumento13 páginasTexto 1 CARACTERISTICAS - BASICAS - DEL - SISTEMA - NERVIOSOjacqueline vallejosAún no hay calificaciones
- Técnicas Quiroprácticas IndirectasDocumento14 páginasTécnicas Quiroprácticas IndirectaspablomyloveAún no hay calificaciones
- Sistema Nervioso Vanessa UniversidadDocumento9 páginasSistema Nervioso Vanessa UniversidadAdriana MateranoAún no hay calificaciones
- NeurofisioDocumento5 páginasNeurofisioMargarita pspAún no hay calificaciones
- Sistema Nervioso, Linfático e Inmunológico, Endócrino y TegumentarioDocumento52 páginasSistema Nervioso, Linfático e Inmunológico, Endócrino y TegumentarioIara DelpardoAún no hay calificaciones
- Neuro - Generalidades Del Sistema Nervioso.Documento48 páginasNeuro - Generalidades Del Sistema Nervioso.hocastro15Aún no hay calificaciones
- El Sistema NerviosoDocumento11 páginasEl Sistema NerviosoAlex EbertAún no hay calificaciones
- Apuntes NeuroanatomíaDocumento109 páginasApuntes NeuroanatomíaMichelle Acuña EspinozaAún no hay calificaciones
- Guia de Lectura Conferencia 31 Sigmund FreudDocumento1 páginaGuia de Lectura Conferencia 31 Sigmund FreudIaia RuizAún no hay calificaciones
- Esquemas Primera ParteDocumento3 páginasEsquemas Primera ParteIaia RuizAún no hay calificaciones
- Estado Nacion, Globalización y DemocraciaDocumento10 páginasEstado Nacion, Globalización y DemocraciaIaia RuizAún no hay calificaciones
- OszlakDocumento12 páginasOszlakIaia Ruiz100% (1)
- Conclusion 1 IntegradorDocumento2 páginasConclusion 1 IntegradorIaia RuizAún no hay calificaciones
- Desigualdades EducativasDocumento4 páginasDesigualdades EducativasIaia RuizAún no hay calificaciones
- Filogenia Del Sistema NerviosoDocumento18 páginasFilogenia Del Sistema NerviosoIaia RuizAún no hay calificaciones
- Coloquio TeoriasDocumento1 páginaColoquio TeoriasIaia RuizAún no hay calificaciones
- Presentación Clase 23 de Agosto 2021Documento9 páginasPresentación Clase 23 de Agosto 2021Iaia RuizAún no hay calificaciones
- Leyes y ResolucionesDocumento1 páginaLeyes y ResolucionesIaia RuizAún no hay calificaciones
- De Giorgi y Plan de EstudiosDocumento13 páginasDe Giorgi y Plan de EstudiosIaia RuizAún no hay calificaciones
- Resumen El Conocimiento Del Profesor Alcala2Documento2 páginasResumen El Conocimiento Del Profesor Alcala2Iaia RuizAún no hay calificaciones
- Teoria GuestaltDocumento11 páginasTeoria GuestaltIaia RuizAún no hay calificaciones
- TEXTO 12. Reportaje A Gardner (Revista Zona Educativa)Documento4 páginasTEXTO 12. Reportaje A Gardner (Revista Zona Educativa)Iaia RuizAún no hay calificaciones
- Sociobiologia - Lazaro y FernandezDocumento31 páginasSociobiologia - Lazaro y FernandezPalmiro Ivan ZambranoAún no hay calificaciones
- Unidad V. Gardner y PerkinsDocumento21 páginasUnidad V. Gardner y PerkinsIaia RuizAún no hay calificaciones
- Aportes para El Desarrollo Curricular Psicologia EducacionalDocumento6 páginasAportes para El Desarrollo Curricular Psicologia EducacionalIaia RuizAún no hay calificaciones
- Plantilla Envio EncomiendaDocumento1 páginaPlantilla Envio EncomiendaIaia RuizAún no hay calificaciones
- TEXTO 6. Teoria de La Gestalt. (Pozo) PDFDocumento6 páginasTEXTO 6. Teoria de La Gestalt. (Pozo) PDFIaia RuizAún no hay calificaciones
- Res 3080 Plan Estratégico Educativo 2020 PDFDocumento4 páginasRes 3080 Plan Estratégico Educativo 2020 PDFIaia Ruiz100% (1)
- TEXTO 6. Teoria de La Gestalt. (Pozo) PDFDocumento6 páginasTEXTO 6. Teoria de La Gestalt. (Pozo) PDFIaia RuizAún no hay calificaciones
- If 2020 01036131 Uba Uba PDFDocumento1 páginaIf 2020 01036131 Uba Uba PDFIaia RuizAún no hay calificaciones
- Guía de Orientación Ciencias Naturales y Educación Ambiental - 5ºDocumento55 páginasGuía de Orientación Ciencias Naturales y Educación Ambiental - 5ºPaula Carrillo0% (1)
- UNIDAD EDUCATIVA Jostin JavierDocumento25 páginasUNIDAD EDUCATIVA Jostin JavierTabian Toala FigueroaAún no hay calificaciones
- CORRELACIóN ESTRATIGRAFICA - OKDocumento15 páginasCORRELACIóN ESTRATIGRAFICA - OKJhonatan Valenzuela H100% (1)
- 3.4 DPRDocumento3 páginas3.4 DPRAlex PerezAún no hay calificaciones
- TunjaDocumento7 páginasTunjaWendy Yuliana Cante León100% (1)
- Examen de Lectura de Cartas y Simbolos - CompressDocumento5 páginasExamen de Lectura de Cartas y Simbolos - CompressRonal FernandezAún no hay calificaciones
- Trabajo Potencia y EnergiaDocumento5 páginasTrabajo Potencia y Energiaandres garciaAún no hay calificaciones
- Proyecto10 - San Juan ÁraDocumento14 páginasProyecto10 - San Juan ÁraCesar OlazarAún no hay calificaciones
- TEMA 1 ANEXO 3 Equipo Botes y Balsas PDFDocumento17 páginasTEMA 1 ANEXO 3 Equipo Botes y Balsas PDFangel33300% (1)
- Experimento Torre de LiquidosDocumento4 páginasExperimento Torre de LiquidosMariaFacebAún no hay calificaciones
- IPERC y Mapa de RiesgosDocumento43 páginasIPERC y Mapa de RiesgosFSS PerúAún no hay calificaciones
- Cartera de Proyectos para EndeudamientoDocumento64 páginasCartera de Proyectos para EndeudamientoOrlando Llaya WongAún no hay calificaciones
- Sistema LagunarDocumento4 páginasSistema LagunarJesus NieblasAún no hay calificaciones
- Protocolo de Bioseguridad - 238 - 2023Documento14 páginasProtocolo de Bioseguridad - 238 - 2023Alejandra Rodriguez ErazoAún no hay calificaciones
- Proyecto Acroleina FINALDocumento29 páginasProyecto Acroleina FINALFiorela Torpoco100% (1)
- Portafolio FerdyyyyDocumento54 páginasPortafolio FerdyyyyWillmer ZaratAún no hay calificaciones
- CUANTI Esquema Muy CompactoDocumento36 páginasCUANTI Esquema Muy CompactoDana MAún no hay calificaciones
- FICHAS - Del 18noviembre1GRADODocumento8 páginasFICHAS - Del 18noviembre1GRADOAracely VillalvaAún no hay calificaciones
- Adgem Rod Ram 2019Documento198 páginasAdgem Rod Ram 2019Silver Huayapa HerreraAún no hay calificaciones
- Memoria Descriptiva General Villa Cristo - YuraDocumento4 páginasMemoria Descriptiva General Villa Cristo - YuraJonathan LeonAún no hay calificaciones
- Comité de Expertos de La OMS en Especificaciones 1Documento14 páginasComité de Expertos de La OMS en Especificaciones 1SalazarValethLeidyAún no hay calificaciones
- S02 - Planeamiento, Programación y Control de La Producción, Productividad e Indicadores de ProducciónDocumento39 páginasS02 - Planeamiento, Programación y Control de La Producción, Productividad e Indicadores de ProducciónJoshua CrederAún no hay calificaciones
- Catalogo de CuentasDocumento13 páginasCatalogo de CuentasGeidi VelezAún no hay calificaciones
- PA3 Consolidado 2 - Circuitos Electricos - 2022 - 20JOSE CARPIODocumento6 páginasPA3 Consolidado 2 - Circuitos Electricos - 2022 - 20JOSE CARPIOEYMI LORENA VILCHEZ RICALDIAún no hay calificaciones
- Reporte Libro - Dejar Ir - Del Dr. David HawkinsDocumento5 páginasReporte Libro - Dejar Ir - Del Dr. David HawkinsMarcia OrozcoAún no hay calificaciones
- 06 Examen Enero Sexto Grado 2020Documento11 páginas06 Examen Enero Sexto Grado 2020Cosme VelascoAún no hay calificaciones
- En Que Estabas PensandoDocumento30 páginasEn Que Estabas PensandoUziel Jordán Martínez LópezAún no hay calificaciones