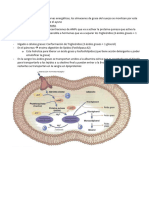
También podría gustarte
- Cómo comprender la fotosíntesis con Max Axiom, supercientíficoDe EverandCómo comprender la fotosíntesis con Max Axiom, supercientíficoCalificación: 4 de 5 estrellas4/5 (2)
- Biología - Química UNED, Tema7Documento3 páginasBiología - Química UNED, Tema7Marta Arias FernandezAún no hay calificaciones
- Fotosíntesis: proceso, organelos, pigmentos y fasesDocumento14 páginasFotosíntesis: proceso, organelos, pigmentos y fasesAngela CVAún no hay calificaciones
- FOTOSINTESIS de Las PlatasDocumento15 páginasFOTOSINTESIS de Las PlatasRuben TorresAún no hay calificaciones
- BC10 FotosintesisDocumento16 páginasBC10 Fotosintesissociedad tapia y bazaes sociedad tapia y bazaesAún no hay calificaciones
- Preguntas Fotosíntesis JhfdtrwqxzsaDocumento16 páginasPreguntas Fotosíntesis JhfdtrwqxzsaNicolee MartinezAún no hay calificaciones
- FOTOSÍNTESIS HPDocumento8 páginasFOTOSÍNTESIS HPPeter P. R. MoralesAún no hay calificaciones
- Fotosintesis y RespiracionDocumento7 páginasFotosintesis y Respiracionemanuel sanchez sotoAún no hay calificaciones
- Practica 4Documento10 páginasPractica 4Yessi HdezAún no hay calificaciones
- Estructura Del Cloroplasto y de Las Mebranas FotosinteticasDocumento4 páginasEstructura Del Cloroplasto y de Las Mebranas Fotosinteticasloky100% (3)
- Resúmenes de FotosíntesisDocumento18 páginasResúmenes de FotosíntesisIsaacAún no hay calificaciones
- FOTOTROFÍADocumento6 páginasFOTOTROFÍAKarin ParraAún no hay calificaciones
- Documento (4)Documento10 páginasDocumento (4)Angie UrecheAún no hay calificaciones
- Formas de Transportar La EnergiaDocumento15 páginasFormas de Transportar La EnergialinaAún no hay calificaciones
- Guía fotosíntesis energía ecosistemaDocumento16 páginasGuía fotosíntesis energía ecosistemajoe videnAún no hay calificaciones
- Fotosintesis 2Documento4 páginasFotosintesis 2nacho cervantesAún no hay calificaciones
- Rincon Del VagoDocumento11 páginasRincon Del VagoBlanca Gallego TévarAún no hay calificaciones
- Obtención de Energía y Síntesis de Compuestos Orgánicos en La Célula VegetalDocumento14 páginasObtención de Energía y Síntesis de Compuestos Orgánicos en La Célula VegetaljenyldaAún no hay calificaciones
- BIOLOGÌADocumento2 páginasBIOLOGÌAMelchor Campos TicllaAún no hay calificaciones
- FOTOSÍNTESISDocumento3 páginasFOTOSÍNTESISCafe CitoAún no hay calificaciones
- Apuntes Bio Parcial 2Documento19 páginasApuntes Bio Parcial 2Jheyris PintoAún no hay calificaciones
- La FotosíntesisDocumento8 páginasLa FotosíntesisSergi Pillaca100% (1)
- FotosintesisDocumento26 páginasFotosintesisDaniel Alexander Echeverry GambaAún no hay calificaciones
- Biol Cap 7 Fotosíntesis PresenDocumento61 páginasBiol Cap 7 Fotosíntesis PresenPriscila RomanAún no hay calificaciones
- La FotosintesisDocumento8 páginasLa Fotosintesistrs21Aún no hay calificaciones
- Semana 4 - Resumen - FotosíntesisDocumento5 páginasSemana 4 - Resumen - FotosíntesisKarlaAún no hay calificaciones
- bloque metabolismoDocumento12 páginasbloque metabolismoYaritzaAún no hay calificaciones
- LA ENERGIA Y SU EVOLUCION EN EL TIEMPO EnsayoDocumento4 páginasLA ENERGIA Y SU EVOLUCION EN EL TIEMPO EnsayojuliAún no hay calificaciones
- Las Fases Luminosas de La Fotosintesis.Documento4 páginasLas Fases Luminosas de La Fotosintesis.Summer LBAún no hay calificaciones
- Tema 7.Documento5 páginasTema 7.Erika Luna JiménezAún no hay calificaciones
- Preguntas Fotosintesis ResueltosDocumento4 páginasPreguntas Fotosintesis ResueltosAmaya Pazdelavega QuirozAún no hay calificaciones
- Los cloroplastos y la fotosíntesisDocumento11 páginasLos cloroplastos y la fotosíntesisDani LoaizaAún no hay calificaciones
- SESIÓN N°11 - BIOLOGÍA - 4° AÑO NsaDocumento2 páginasSESIÓN N°11 - BIOLOGÍA - 4° AÑO NsaKarina NeyraAún no hay calificaciones
- Guia 6Documento4 páginasGuia 6Andrea JJJAún no hay calificaciones
- Proceso fotosíntesisDocumento3 páginasProceso fotosíntesisIvan De la RosaAún no hay calificaciones
- Fotosíntesis Bacteriana PDFDocumento17 páginasFotosíntesis Bacteriana PDFCristhian NcAún no hay calificaciones
- ANABOLISMODocumento7 páginasANABOLISMOcarmenzg30Aún no hay calificaciones
- CloroplastosDocumento5 páginasCloroplastosrobertojrezAún no hay calificaciones
- Cuáles Son Las Etapas de La FotosíntesisDocumento4 páginasCuáles Son Las Etapas de La FotosíntesisLuis The GamerAún no hay calificaciones
- 28 Solucionario FotosíntesisDocumento16 páginas28 Solucionario FotosíntesisEvelyn Muñoz SotoAún no hay calificaciones
- Taller FotosíntesisDocumento10 páginasTaller Fotosíntesisdeivy alvarezAún no hay calificaciones
- Anabolismo IDocumento8 páginasAnabolismo IClaudia Lopez VicenteAún no hay calificaciones
- Nutricion Autotrofa Nuevo Documento de Microsoft Office WordDocumento11 páginasNutricion Autotrofa Nuevo Documento de Microsoft Office WordLisseth Aguero AbarcaAún no hay calificaciones
- Practica 1 DesarrolloDocumento8 páginasPractica 1 DesarrolloThaily RiosAún no hay calificaciones
- Fotofosforilacin Ciclica y No CclicaDocumento43 páginasFotofosforilacin Ciclica y No CclicaLuis GomezAún no hay calificaciones
- Lectura - AnabolismoDocumento9 páginasLectura - AnabolismoSegundo FernandezAún no hay calificaciones
- Preguntas Fotosintesis Guia de EstudioDocumento8 páginasPreguntas Fotosintesis Guia de Estudiocarolinalisseth9030velezAún no hay calificaciones
- Proceso metabólico por el cual las plantas y algas transforman la energía luminosa en energía químicaDocumento4 páginasProceso metabólico por el cual las plantas y algas transforman la energía luminosa en energía químicaJackson López LaviAún no hay calificaciones
- Fase luminosa fotosíntesis clavesDocumento3 páginasFase luminosa fotosíntesis clavesDaniel GarciaAún no hay calificaciones
- Etapas de La FotosíntesisDocumento12 páginasEtapas de La FotosíntesisASHLEY SOTOAún no hay calificaciones
- UNIDAD 2 Temas 13 y 14Documento38 páginasUNIDAD 2 Temas 13 y 14LuisAún no hay calificaciones
- Una Forma Más General de Interpretar Las Reacciones ÓxidoDocumento3 páginasUna Forma Más General de Interpretar Las Reacciones ÓxidoAlex Hdez AlfaroAún no hay calificaciones
- FotosintesisDocumento4 páginasFotosintesismelisalosiggioAún no hay calificaciones
- Orientaciones:: Castilla-León / Junio 98. Cou / Biología / Célula Y Sus Componentes. / Opción A / #3Documento3 páginasOrientaciones:: Castilla-León / Junio 98. Cou / Biología / Célula Y Sus Componentes. / Opción A / #3lauramatesAún no hay calificaciones
- Fotosintesis y Como Se ClasificanDocumento8 páginasFotosintesis y Como Se ClasificanMario Guzman MartinezAún no hay calificaciones
- Fotosíntesis: Proceso vital que convierte la energía solar en energía químicaDocumento45 páginasFotosíntesis: Proceso vital que convierte la energía solar en energía químicaMarcos ChávezAún no hay calificaciones
- Fotosintesis y La Respiracion Lab de BiologiaDocumento4 páginasFotosintesis y La Respiracion Lab de BiologiaDaniela Sánchez MedinaAún no hay calificaciones
- Cadena Transportadora de ElectronesDocumento8 páginasCadena Transportadora de Electroneszulycastillo_2496Aún no hay calificaciones
- Problemas de GenéticaDocumento8 páginasProblemas de GenéticaSofia CasasAún no hay calificaciones
- DiferenciasC3C4plantasDocumento5 páginasDiferenciasC3C4plantasMARIANA MEJIA VERGARAAún no hay calificaciones
- Trabajo de Catabolismo y AnabolismoDocumento6 páginasTrabajo de Catabolismo y AnabolismoYojana PerezAún no hay calificaciones
- Docente:: Saldaña Rojas, Edinson Nicolas Curso: Bioquímica de AlimentosDocumento22 páginasDocente:: Saldaña Rojas, Edinson Nicolas Curso: Bioquímica de AlimentosMAYRA LIZETH MORI HOYOSAún no hay calificaciones
- BMC Fermentacion RuminalDocumento25 páginasBMC Fermentacion Ruminalapi-3743425100% (1)
- Resumen MetabolismoDocumento9 páginasResumen MetabolismoFer RiveraAún no hay calificaciones
- GluconeogenesisDocumento2 páginasGluconeogenesisAnnyVal RMAún no hay calificaciones
- UNIBAR-BIOQUIMICA-NUCLEOTIDOSDocumento4 páginasUNIBAR-BIOQUIMICA-NUCLEOTIDOSPrime VideoAún no hay calificaciones
- CloroplastosDocumento5 páginasCloroplastosMario Lagunes NegroAún no hay calificaciones
- Ruta MetabólicaDocumento20 páginasRuta MetabólicaAna Milena Riascos GrajalesAún no hay calificaciones
- Problemas de Catálisis Química PDFDocumento9 páginasProblemas de Catálisis Química PDFnaleny ruiz cruzAún no hay calificaciones
- Respiración en PlantasDocumento16 páginasRespiración en PlantasccasalvaAún no hay calificaciones
- MITOCONDRIA ExposicionDocumento30 páginasMITOCONDRIA ExposicionKaren Vigo0% (1)
- Tríptico de La Glucolisis.Documento2 páginasTríptico de La Glucolisis.Maritza Ramírez ChanAún no hay calificaciones
- Adme LípidosDocumento85 páginasAdme LípidosCarolina ZegarraAún no hay calificaciones
- Tecnicas para El Estudio de La Funcion MitocondrialDocumento2 páginasTecnicas para El Estudio de La Funcion MitocondrialFer MartellAún no hay calificaciones
- Metabolismo de Compuestos NitrogenadosDocumento13 páginasMetabolismo de Compuestos NitrogenadosManuel NinahuancaAún no hay calificaciones
- Presentaciòn Metabolismo Monog y Rumiantes Resumen UnillanosDocumento17 páginasPresentaciòn Metabolismo Monog y Rumiantes Resumen UnillanosCINDY CAROLINA JIMENEZ GILAún no hay calificaciones
- AnabolismoDocumento5 páginasAnabolismoDAYANA CARHUAPOMA CANALESAún no hay calificaciones
- Guarda tu progresoDocumento6 páginasGuarda tu progresoMartrem81Aún no hay calificaciones
- FERMENTACIÓNDocumento3 páginasFERMENTACIÓNAngie BravoAún no hay calificaciones
- ADIEL RIVAS-GUÍA DE TRABAJO No. 5 Bioenergética Del Metabolismo de Los CarbohidratosDocumento3 páginasADIEL RIVAS-GUÍA DE TRABAJO No. 5 Bioenergética Del Metabolismo de Los CarbohidratosRosalina Perez QuisqueAún no hay calificaciones
- 7 Acidosis-MetablicaDocumento29 páginas7 Acidosis-MetablicaVictor Cercado VásquezAún no hay calificaciones
- Modelos bioquímicos: La glucólisisDocumento6 páginasModelos bioquímicos: La glucólisisNatalia TorresAún no hay calificaciones
- Respiración Celular AerobicaDocumento6 páginasRespiración Celular AerobicaKoby RamosAún no hay calificaciones
- Metabolismo de alimentos y formación de compuestos corporalesDocumento30 páginasMetabolismo de alimentos y formación de compuestos corporalesliliana martinez0% (1)
- GluconeognesisDocumento38 páginasGluconeognesisDiego DGAún no hay calificaciones
- Metabolismo de Los Hidratos de CarbonoDocumento21 páginasMetabolismo de Los Hidratos de CarbonoJorge CopelAún no hay calificaciones
- Anabolismo de LípidosDocumento17 páginasAnabolismo de LípidosAngie YáñezAún no hay calificaciones
- CUESTIONARIODocumento3 páginasCUESTIONARIOalexaAún no hay calificaciones
- Beta OxidaciónDocumento3 páginasBeta OxidaciónSofía DevittaAún no hay calificaciones