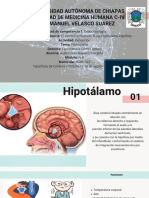
También podría gustarte
- Endogrionologia y Sexualidad 26-05-22Documento21 páginasEndogrionologia y Sexualidad 26-05-22Oswaldo AlvaradoAún no hay calificaciones
- Endocrino 1Documento8 páginasEndocrino 1Alejandra SaizaAún no hay calificaciones
- HIPOFISIS Monografia Trabajo Final REPARTIDODocumento14 páginasHIPOFISIS Monografia Trabajo Final REPARTIDOSebastian Florentino VereusAún no hay calificaciones
- Hormonas NeurohipofisiariasDocumento11 páginasHormonas NeurohipofisiariasMiguel EstradaAún no hay calificaciones
- NEUROENDOCRINOLOGÍADocumento31 páginasNEUROENDOCRINOLOGÍAsantiagolunarevalo0% (1)
- Presentacion HipofisisDocumento41 páginasPresentacion HipofisisViictoor Roobeerto Goonzaalez SaancheezAún no hay calificaciones
- EQUIPO Hipotálamo - 1Documento59 páginasEQUIPO Hipotálamo - 1Emmanuel HernandezAún no hay calificaciones
- HipotalamoDocumento8 páginasHipotalamoJosé MalaspinaAún no hay calificaciones
- Clase Histologia ENDOCRINODocumento69 páginasClase Histologia ENDOCRINOValeria NoireAún no hay calificaciones
- NeurohipófisisDocumento36 páginasNeurohipófisisIsabella Hoyos MaderaAún no hay calificaciones
- Berne y Levy. Fisiología Neurohipófisis PDFDocumento19 páginasBerne y Levy. Fisiología Neurohipófisis PDFAngel Dosantos AveiroAún no hay calificaciones
- La HipofisisDocumento14 páginasLa HipofisisTatianaCruzAún no hay calificaciones
- Anatomía 1Documento2 páginasAnatomía 1Moises RodriguezAún no hay calificaciones
- 16 Sistema EndocrinoDocumento12 páginas16 Sistema EndocrinoSer leAún no hay calificaciones
- Sistema - Endocrino (1) PPTDocumento54 páginasSistema - Endocrino (1) PPTJefferson CollachaguaAún no hay calificaciones
- Seminario HipotálamoDocumento20 páginasSeminario HipotálamoValeria CárdenasAún no hay calificaciones
- Guia 1 Endocrino Eje Hipotalamo Hipofisis PDFDocumento5 páginasGuia 1 Endocrino Eje Hipotalamo Hipofisis PDFbefamaAún no hay calificaciones
- Sistema EndocrinoDocumento34 páginasSistema EndocrinoAzula91100% (1)
- Hipotálamo (1Documento16 páginasHipotálamo (1nayeli esmeralda avalos celisAún no hay calificaciones
- Principios de EndocrinologíaDocumento52 páginasPrincipios de EndocrinologíadianaAún no hay calificaciones
- HIPOFISISDocumento5 páginasHIPOFISISRobin WelchAún no hay calificaciones
- Anatomia HH - Salvador SilvaDocumento2 páginasAnatomia HH - Salvador SilvaDaniela SalvadorAún no hay calificaciones
- Hipotálamo - Hipofisis TrastornosDocumento83 páginasHipotálamo - Hipofisis TrastornosJhon GregoriAún no hay calificaciones
- Anatomia Salas PDFDocumento28 páginasAnatomia Salas PDFMarita TrujilloAún no hay calificaciones
- Lab2 EndoDocumento11 páginasLab2 EndoJoshua MenesesAún no hay calificaciones
- Guia ENDOCRINODocumento8 páginasGuia ENDOCRINOGabriel IbarraAún no hay calificaciones
- NeurohipófisisDocumento2 páginasNeurohipófisisTwicface ツAún no hay calificaciones
- 08 - Tema Endocrino 1 - Hipotálamo-Hipófisis-Guía Práctica-Histología Sistemática y Embriología-2021Documento13 páginas08 - Tema Endocrino 1 - Hipotálamo-Hipófisis-Guía Práctica-Histología Sistemática y Embriología-2021Alejandra GonzálezAún no hay calificaciones
- HIPOTALAMODocumento1 páginaHIPOTALAMOMirlaAún no hay calificaciones
- Sistema EndocrinoDocumento30 páginasSistema EndocrinobetosalAún no hay calificaciones
- HipotálamoDocumento45 páginasHipotálamoLuis BatistaAún no hay calificaciones
- Sistema Nervioso 6oDocumento34 páginasSistema Nervioso 6oJulián Jaber GuaidaAún no hay calificaciones
- Regulacion Hipotalamica de Las Funciones HormonalesDocumento10 páginasRegulacion Hipotalamica de Las Funciones HormonalesMitzi Kimberly Torres AlanizAún no hay calificaciones
- Histología de La HipofisisDocumento13 páginasHistología de La HipofisisLucia Saavedra LozadaAún no hay calificaciones
- TRIPTICODocumento2 páginasTRIPTICOAnonymous cY6MO9uF6PAún no hay calificaciones
- Organos EndocrinosDocumento6 páginasOrganos EndocrinosAioros SagitarioAún no hay calificaciones
- El HipotálamoDocumento12 páginasEl HipotálamoMarce MartinezAún no hay calificaciones
- Wa0088Documento17 páginasWa0088Yulissa Conca MendozaAún no hay calificaciones
- Histología Del Sistema EndocrinoDocumento27 páginasHistología Del Sistema EndocrinoMariana BorjaAún no hay calificaciones
- Eje HipotálamoDocumento8 páginasEje HipotálamoFede AranaAún no hay calificaciones
- Hipotálamo y Glándula Hipófisis (Libro)Documento6 páginasHipotálamo y Glándula Hipófisis (Libro)Erika LoyarteAún no hay calificaciones
- Estructura Interna Del Hemisferio CerebralDocumento83 páginasEstructura Interna Del Hemisferio CerebraldaniinocomeAún no hay calificaciones
- Los Órganos Circunventriculares No Disponen de Barrera HematoencefálicaDocumento10 páginasLos Órganos Circunventriculares No Disponen de Barrera HematoencefálicaAlba López NúñezAún no hay calificaciones
- Presentacion Anatomia Humana Medicina Con Diagramas Bonita Colores PastelDocumento10 páginasPresentacion Anatomia Humana Medicina Con Diagramas Bonita Colores PastelJean Coronel MelendresAún no hay calificaciones
- Sistema Endocrino22Documento142 páginasSistema Endocrino22Monse AlfaroAún no hay calificaciones
- Anatomía Del HipotálamoDocumento7 páginasAnatomía Del HipotálamoRaymi Luis Ramirez100% (12)
- OrexinasDocumento6 páginasOrexinasAngelAún no hay calificaciones
- 14 HipotálamoDocumento22 páginas14 HipotálamoJheisonAún no hay calificaciones
- HipofisisDocumento7 páginasHipofisisRosario CaleroAún no hay calificaciones
- Universidad de Las Ciencias de La Salud - HipofisisDocumento31 páginasUniversidad de Las Ciencias de La Salud - Hipofisismarlon urbanejaAún no hay calificaciones
- Sistema Nervioso Central y PeriféricoDocumento31 páginasSistema Nervioso Central y PeriféricoGabriela QuichimboAún no hay calificaciones
- SEMANA 8 - Sistema EndocrinoDocumento14 páginasSEMANA 8 - Sistema Endocrinoamuven1967Aún no hay calificaciones
- Teoria Sistema NeuroendocrinoDocumento9 páginasTeoria Sistema NeuroendocrinoBRENDA ELIZABETH QUISPE QUISPEAún no hay calificaciones
- 2 Hipotálamo y AdenohipófisisDocumento11 páginas2 Hipotálamo y AdenohipófisisBarbara VegaAún no hay calificaciones
- 3 y 4 HipofisisDocumento6 páginas3 y 4 HipofisisAníbal CIAún no hay calificaciones
- Hipofisis HistologiaDocumento6 páginasHipofisis HistologiaAníbal CIAún no hay calificaciones
- HIPOTALAMODocumento30 páginasHIPOTALAMORafael Falcon AriasAún no hay calificaciones
- El cannabis en patologías del sistema nervioso centralDe EverandEl cannabis en patologías del sistema nervioso centralAún no hay calificaciones
- Anatomía y Fisiología del Sistema NerviosoDe EverandAnatomía y Fisiología del Sistema NerviosoCalificación: 4 de 5 estrellas4/5 (19)
- Anatomia y Fisiología del Sistema Nervioso IIDe EverandAnatomia y Fisiología del Sistema Nervioso IICalificación: 4 de 5 estrellas4/5 (28)
- Pasos para Tomar La Presión ArterialDocumento2 páginasPasos para Tomar La Presión ArterialSonia Ortiz DominguezAún no hay calificaciones
- Activación y Disfunción Endotelial en Síndrome Metabólico, Diabetes Tipo 2 y Enfermedad Por Coronavirus 2019Documento22 páginasActivación y Disfunción Endotelial en Síndrome Metabólico, Diabetes Tipo 2 y Enfermedad Por Coronavirus 2019Alejandro Londoño AmarilesAún no hay calificaciones
- AcuaporinasDocumento8 páginasAcuaporinasFabiola MartinezAún no hay calificaciones
- Pro - Sarampion y RubeolaDocumento22 páginasPro - Sarampion y RubeolaBrando Peñaranda RodriguezAún no hay calificaciones
- PRO ChikunguñaDocumento17 páginasPRO ChikunguñaZuley QuirogaAún no hay calificaciones
- 1º Cien Nat PC e 28 Setiembre 210928 081747Documento5 páginas1º Cien Nat PC e 28 Setiembre 210928 081747Ever Medina GonzálezAún no hay calificaciones
- El Aprendizaje Visto Con Un Enfoque NeurocientíficoDocumento6 páginasEl Aprendizaje Visto Con Un Enfoque NeurocientíficoAndrea MartinezAún no hay calificaciones
- Neoformaciones Frecuentes de La ConjuntivaDocumento10 páginasNeoformaciones Frecuentes de La ConjuntivaDaniela Ruiz MartinezAún no hay calificaciones
- Manual Instalacion JenkinsDocumento7 páginasManual Instalacion JenkinsfredyAún no hay calificaciones
- Gran Memorial Del Conde Duque de OlivaresDocumento4 páginasGran Memorial Del Conde Duque de Olivaresrapevar100% (1)
- Mi CuerpoDocumento30 páginasMi CuerpoDarly Yohana50% (2)
- Curso de RedacciónDocumento11 páginasCurso de RedacciónBrilly CorreaAún no hay calificaciones
- Lectura Y Escritura de Textos: Una Doble Competencia para Estudiantes Universitarios 1. Estrategias GeneralesDocumento4 páginasLectura Y Escritura de Textos: Una Doble Competencia para Estudiantes Universitarios 1. Estrategias GeneralesEdgar Campaña20% (5)
- El Concepto Hayekiano de Orden Espontáneo. Federico Sosa Valle.Documento17 páginasEl Concepto Hayekiano de Orden Espontáneo. Federico Sosa Valle.StuttgartAún no hay calificaciones
- GUÍA No 2 - Medidas de Tendencia Central y Dispersión - RRR PDFDocumento6 páginasGUÍA No 2 - Medidas de Tendencia Central y Dispersión - RRR PDFOrlando K'rmona Martinez100% (1)
- ¿Quién Era El Joven Que Seguía Desnudo A Jesús Blog de Francisco Díaz SJDocumento4 páginas¿Quién Era El Joven Que Seguía Desnudo A Jesús Blog de Francisco Díaz SJHARVEY ALEXANDER SUAREZ MARTINEZAún no hay calificaciones
- Clinica PenalDocumento7 páginasClinica PenalGustavo CastañedaAún no hay calificaciones
- El Entorno de La AlabanzaDocumento3 páginasEl Entorno de La AlabanzamanueladrianaAún no hay calificaciones
- 2.2 Tipos de SoldaduraDocumento11 páginas2.2 Tipos de SoldaduraHebander EsauAún no hay calificaciones
- La Formación Del Lector Literario Teresa ColomerDocumento15 páginasLa Formación Del Lector Literario Teresa ColomerAída De Paula100% (1)
- KWE SX WET ÛSKIWE NXI ÎTXISA Pedagogía de La Cultura ActivaDocumento161 páginasKWE SX WET ÛSKIWE NXI ÎTXISA Pedagogía de La Cultura ActivaMauricio BarreraAún no hay calificaciones
- Cuadro Sinóptico e Informe Sobre Los CostosDocumento7 páginasCuadro Sinóptico e Informe Sobre Los CostosEstefania R.cAún no hay calificaciones
- Desarrollo de Una Aplicación Móvil Multiplataforma de Mensajería Instantanea para Agentes EmpresarialesDocumento103 páginasDesarrollo de Una Aplicación Móvil Multiplataforma de Mensajería Instantanea para Agentes EmpresarialesJose FranciscoAún no hay calificaciones
- Mousse de Sangrecita TripticoDocumento2 páginasMousse de Sangrecita TripticoEdward RosadoAún no hay calificaciones
- Formato PasantiasDocumento25 páginasFormato PasantiasLibrosCristianosAún no hay calificaciones
- Cuidado de AparatosDocumento2 páginasCuidado de AparatosLucia sales estevezAún no hay calificaciones
- Fuente ConmutadaDocumento33 páginasFuente ConmutadaRoberto Iván M BAún no hay calificaciones
- Clasificacion Del Gasto PublicoDocumento6 páginasClasificacion Del Gasto PublicoEyeline Aguilar100% (1)
- Minuta Jesus Froilan Ramos Coaquira MinutaaDocumento10 páginasMinuta Jesus Froilan Ramos Coaquira MinutaaEH LeynaAún no hay calificaciones
- Tarea Algebra N 6Documento1 páginaTarea Algebra N 6Henry Condori HuallpaAún no hay calificaciones
- ROMANO Braudel Nosotros Los DemasDocumento14 páginasROMANO Braudel Nosotros Los DemasDarío HerreraAún no hay calificaciones
- "Evolución de Las Funciones de La Educación Física": Issn 1988-6047 DEP. LEGAL: GR 2922/2007Documento9 páginas"Evolución de Las Funciones de La Educación Física": Issn 1988-6047 DEP. LEGAL: GR 2922/2007Diego Gálvez GimenoAún no hay calificaciones
- 1.6 Transversal y LongitudinalDocumento10 páginas1.6 Transversal y LongitudinalJavier Lara Flores0% (1)
- Hemorragia CerebralDocumento29 páginasHemorragia CerebralRuth Aseret Gallegos PerroniAún no hay calificaciones
- Funcion Operacional en Las OrganizacionesDocumento4 páginasFuncion Operacional en Las OrganizacionesJazmin AlejandraAún no hay calificaciones