También podría gustarte
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017De EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Aún no hay calificaciones
- Bioenergética. Introducción a la teoría quimiosmóticaDe EverandBioenergética. Introducción a la teoría quimiosmóticaAún no hay calificaciones
- NeuroreceptoresDocumento4 páginasNeuroreceptoresFlor de LotoAún no hay calificaciones
- Reporte Ing GenéticaDocumento8 páginasReporte Ing GenéticaMike BurtonAún no hay calificaciones
- Seminario 1BDocumento10 páginasSeminario 1BRomina NogueiraAún no hay calificaciones
- Análisis Del Dna Plásmídico de Escherichia Coli Transformada Mediante Digestión ConDocumento7 páginasAnálisis Del Dna Plásmídico de Escherichia Coli Transformada Mediante Digestión ConLaura C. CastellanosAún no hay calificaciones
- 02.AB Materiales y MetodosDocumento34 páginas02.AB Materiales y MetodosJennifer Alondra Hernandez ZuñigaAún no hay calificaciones
- Uso de siRNA Sobre El Receptor A Interleucina 7Documento9 páginasUso de siRNA Sobre El Receptor A Interleucina 7Edgardo Becerra BecerraAún no hay calificaciones
- Artículo Final - Equipo 2Documento5 páginasArtículo Final - Equipo 2FRIDA SOFIA GARCIA BARRERAAún no hay calificaciones
- Preparación de CromatinaDocumento6 páginasPreparación de CromatinaNatalia Andrea Nuñez ToledoAún no hay calificaciones
- La Fosforilación Del Núcleo de La Histona Regula La Accesibilidad Del AdnDocumento5 páginasLa Fosforilación Del Núcleo de La Histona Regula La Accesibilidad Del AdneverAún no hay calificaciones
- Practica 11 Perfil Hepatico - Coléstesis-CitólisisDocumento7 páginasPractica 11 Perfil Hepatico - Coléstesis-CitólisisValeria Vargas AguilarAún no hay calificaciones
- Guía de Laboratorio de PCR y ElectroforesisDocumento6 páginasGuía de Laboratorio de PCR y ElectroforesisObleas Pedro A San CarlosAún no hay calificaciones
- Ribosomal FrameshiftingDocumento37 páginasRibosomal FrameshiftingErika CoronadoAún no hay calificaciones
- Efectos Múltiples de La 4 AminopiridinaDocumento5 páginasEfectos Múltiples de La 4 AminopiridinayobanyAún no hay calificaciones
- 14UAQPerezGutierrez-CANALES DE CLORODocumento4 páginas14UAQPerezGutierrez-CANALES DE CLOROChoffiz D Kaulitz Schafer ListingAún no hay calificaciones
- Farmacologia Del Sistema Nervioso Autonomo Parasimpatico-5b26c197d8520Documento11 páginasFarmacologia Del Sistema Nervioso Autonomo Parasimpatico-5b26c197d8520javiera_chingAún no hay calificaciones
- Sinapsis I-2-4Documento59 páginasSinapsis I-2-4Monica RodriguezAún no hay calificaciones
- Memoria Ingenieria GenéticaDocumento9 páginasMemoria Ingenieria GenéticaJavier Gómez PiñeroAún no hay calificaciones
- Traduccion y Sintesis de ProteinasDocumento36 páginasTraduccion y Sintesis de ProteinasSara María Vargas CastilloAún no hay calificaciones
- Casos 2Documento5 páginasCasos 2Mariana NóbregaAún no hay calificaciones
- PCR y ElectroforessisDocumento9 páginasPCR y ElectroforessisLina González GalvánAún no hay calificaciones
- Sintesis Del ArticuloDocumento5 páginasSintesis Del ArticuloGeysiAún no hay calificaciones
- Aislamiento, Caracterización Electroforética y Amplificación Mediante PCR de Dna PlasmídicoDocumento5 páginasAislamiento, Caracterización Electroforética y Amplificación Mediante PCR de Dna PlasmídicoPaola FarfanAún no hay calificaciones
- Electroforesis de DNA Plasmídico de Escherichia Coli en Gel de AgarosaDocumento9 páginasElectroforesis de DNA Plasmídico de Escherichia Coli en Gel de AgarosaFlorencia GobboAún no hay calificaciones
- Guía de Laboratorio de PCR y Electroforesis 2023Documento6 páginasGuía de Laboratorio de PCR y Electroforesis 2023ANDRES FELIPE CAMACHO FIRIGUAAún no hay calificaciones
- Práctica Polimorfismos de Los Genes ABCB1Documento5 páginasPráctica Polimorfismos de Los Genes ABCB1Daniel SchAún no hay calificaciones
- Introducción de La Técnica PCRDocumento6 páginasIntroducción de La Técnica PCRAngie CeronAún no hay calificaciones
- Purificación y Caracterización de Bineatobina Uan Protein Coagulante de Veneno de Serpiente PeruanaDocumento10 páginasPurificación y Caracterización de Bineatobina Uan Protein Coagulante de Veneno de Serpiente PeruanaGustavo RuizAún no hay calificaciones
- Fisiologia RETINADocumento43 páginasFisiologia RETINAAmacilis Suarez100% (1)
- Reporte 1Documento6 páginasReporte 1DenisseAún no hay calificaciones
- Caracterización Molecular y Bioquímica de Una Nueva XilanasaDocumento33 páginasCaracterización Molecular y Bioquímica de Una Nueva XilanasaDiana RodríguezAún no hay calificaciones
- Ligacion Clonacion y Extraccion Del PlasmidoDocumento5 páginasLigacion Clonacion y Extraccion Del PlasmidoManuel Andres Ampuero MartinezAún no hay calificaciones
- TP N 7 Operon Lac Giudici AbrilDocumento8 páginasTP N 7 Operon Lac Giudici AbrilAbril GiudiciAún no hay calificaciones
- J A PINA Practicas de Biologia MolecularDocumento15 páginasJ A PINA Practicas de Biologia MolecularTeresa MontesAún no hay calificaciones
- 23-24 - Bioquimica General Resultados - 090923Documento8 páginas23-24 - Bioquimica General Resultados - 090923Venga ChavalAún no hay calificaciones
- 5to Informe PCRDocumento8 páginas5to Informe PCRKaren Esteves ZúñigaAún no hay calificaciones
- Diseño Experimental Y Análisis de Datos en La Técnica de La QRT-PCR (Quantitive Real Time-Polymerasa Chain Reaction)Documento15 páginasDiseño Experimental Y Análisis de Datos en La Técnica de La QRT-PCR (Quantitive Real Time-Polymerasa Chain Reaction)David OlascoagaAún no hay calificaciones
- Laboratorio Polimorfismo Factor V Leiden Por PCR-RFLPDocumento5 páginasLaboratorio Polimorfismo Factor V Leiden Por PCR-RFLPluxitocoli100% (1)
- EJEMPLO 2 Practica-2-Metodo-De-SangerDocumento4 páginasEJEMPLO 2 Practica-2-Metodo-De-SangerAndy AldamaAún no hay calificaciones
- Pro4 Reg9 ckmb2Documento4 páginasPro4 Reg9 ckmb2Vinicio Contreras CastroAún no hay calificaciones
- Poster Científico LEANGELANDRODocumento5 páginasPoster Científico LEANGELANDROAngela Vanessa Pacheco Flores de ValgasAún no hay calificaciones
- Enzimas de RestriccionDocumento9 páginasEnzimas de RestriccionFRIDA SOFIA GARCIA BARRERAAún no hay calificaciones
- Tecnicas para El Estudio de La Funcion MitocondrialDocumento2 páginasTecnicas para El Estudio de La Funcion MitocondrialFer MartellAún no hay calificaciones
- Síndrome Letal Overo BlancoDocumento32 páginasSíndrome Letal Overo BlancocasanchezisAún no hay calificaciones
- Memorias Escoto AMIDIQ 2012Documento6 páginasMemorias Escoto AMIDIQ 2012Dorita RomanAún no hay calificaciones
- Reaccion en Cadena de La PolimerasaDocumento13 páginasReaccion en Cadena de La PolimerasaGina M LópezAún no hay calificaciones
- GATOLATEDocumento6 páginasGATOLATEnicositja_vangoh91Aún no hay calificaciones
- TESIS VimpresaDocumento6 páginasTESIS VimpresaJose David VelandiaAún no hay calificaciones
- Ensayo Basado en Cuentas para El Control Espacio-Temporal de La Expresión Génica en Sistemas de Transcripción-Traducción Sin CélulasDocumento9 páginasEnsayo Basado en Cuentas para El Control Espacio-Temporal de La Expresión Génica en Sistemas de Transcripción-Traducción Sin CélulasCristian Camilo Ramirez RangelAún no hay calificaciones
- Extracción y Cuantificación de Ácidos NucleicosDocumento6 páginasExtracción y Cuantificación de Ácidos NucleicosLucero Odar CastilloAún no hay calificaciones
- PDF DocumentoDocumento6 páginasPDF DocumentoXdjaja GraciasAún no hay calificaciones
- Prob. Met. 2QxEstudiantes CAST x13x14xDocumento10 páginasProb. Met. 2QxEstudiantes CAST x13x14xarisale1Aún no hay calificaciones
- Artículo - Hidroxilamina Como Agente Mutagénico para Neurospora Crassa Con Corr 1Documento5 páginasArtículo - Hidroxilamina Como Agente Mutagénico para Neurospora Crassa Con Corr 1leilaniAún no hay calificaciones
- Calb Enzima PDFDocumento15 páginasCalb Enzima PDFJuan OlivaAún no hay calificaciones
- TRABAJO PRÁCTICO N°4 y 5 ADN 2022Documento5 páginasTRABAJO PRÁCTICO N°4 y 5 ADN 2022TrinidadAún no hay calificaciones
- MOR y Corteza VisualDocumento5 páginasMOR y Corteza VisualMauricio SaldivarAún no hay calificaciones
- Resumen Sinaptofisina y GlutamatoDocumento4 páginasResumen Sinaptofisina y GlutamatoAndrea Fernanda Vélez ZambranoAún no hay calificaciones
- Protocolo de Extracción de DNA Por Salting-OutDocumento4 páginasProtocolo de Extracción de DNA Por Salting-OutDanny R. MorenoAún no hay calificaciones
- Manual de electrocardiografía canina para estudiantes de medicina veterinariaDe EverandManual de electrocardiografía canina para estudiantes de medicina veterinariaAún no hay calificaciones
- Resumen de Salidroside Atenúa La Neuroinflamación Inducida Por Lipopolisacáridos y El Deterioro Cognitivo en Ratones Con Encefalopatía SépticaDocumento2 páginasResumen de Salidroside Atenúa La Neuroinflamación Inducida Por Lipopolisacáridos y El Deterioro Cognitivo en Ratones Con Encefalopatía SépticaAndrea Fernanda Vélez ZambranoAún no hay calificaciones
- Neuropatía y GABAADocumento4 páginasNeuropatía y GABAAAndrea Fernanda Vélez ZambranoAún no hay calificaciones
- Resumen de Modulación Astrocítica de La Excitabilidad Neuronal A Través de La Amortiguación Espacial K+Documento3 páginasResumen de Modulación Astrocítica de La Excitabilidad Neuronal A Través de La Amortiguación Espacial K+Andrea Fernanda Vélez ZambranoAún no hay calificaciones
- Resumen Sinaptofisina y GlutamatoDocumento4 páginasResumen Sinaptofisina y GlutamatoAndrea Fernanda Vélez ZambranoAún no hay calificaciones
- Marco Teorico SiiiiDocumento68 páginasMarco Teorico Siiiijuan pabloAún no hay calificaciones
- Cuadro Resumen Aparato Rep Femenino y MasculinoDocumento2 páginasCuadro Resumen Aparato Rep Femenino y MasculinoRobbie RivasAún no hay calificaciones
- No Quiero Ir Al PsicologoDocumento2 páginasNo Quiero Ir Al PsicologoPix AyalaAún no hay calificaciones
- Ejercicios para La Columna LumbarDocumento6 páginasEjercicios para La Columna LumbarLupibo PbAún no hay calificaciones
- Libre TaDocumento10 páginasLibre TaAlexanderValenciaRamosAún no hay calificaciones
- Principales Descubrimientos de Craig VenterDocumento3 páginasPrincipales Descubrimientos de Craig VenterNatasha GuilcapiAún no hay calificaciones
- Guía de Recuperacion Grado 6Documento10 páginasGuía de Recuperacion Grado 6Cristian CamiloAún no hay calificaciones
- TALLERDocumento2 páginasTALLERGiovani Albarracín100% (2)
- Olericultura Corregida 2021-IIDocumento207 páginasOlericultura Corregida 2021-IIRocio Irene Lucano TarrilloAún no hay calificaciones
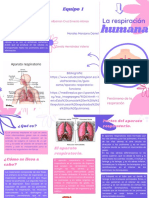
- La Respiracion Humana.Documento2 páginasLa Respiracion Humana.Zavala Hernández Valeria.1IM18Aún no hay calificaciones
- Uso de Pasto Vetiver en Obras de BioingenieríaDocumento129 páginasUso de Pasto Vetiver en Obras de Bioingenieríamarj100000100% (1)
- Reporte Práctica Biología - 3Documento7 páginasReporte Práctica Biología - 3Sara AguirreAún no hay calificaciones
- PLACEDocumento6 páginasPLACEAbdiel Valencia100% (1)
- Pruebas de EnsayoDocumento13 páginasPruebas de EnsayoBaucorAún no hay calificaciones
- Elaboracion de Pastabde Chile HabaneroDocumento8 páginasElaboracion de Pastabde Chile HabaneroGuillermina GutierrezAún no hay calificaciones
- Cuantificación de Microorganismos Por El Método de Recuento en Placa y Técnica Del Número Más ProbableDocumento5 páginasCuantificación de Microorganismos Por El Método de Recuento en Placa y Técnica Del Número Más ProbableJULIAN ANDRES SOTO SANTEROAún no hay calificaciones
- Copia de HTP (Casa - Arbol - Persona)Documento15 páginasCopia de HTP (Casa - Arbol - Persona)fercho xaviAún no hay calificaciones
- 638 CiencioramaDocumento11 páginas638 CiencioramaOscar CortezAún no hay calificaciones
- Aporte de Científicos A La BiologíaDocumento7 páginasAporte de Científicos A La BiologíaHebteco ShadAún no hay calificaciones
- Biología - Repaso Extremo 1Documento7 páginasBiología - Repaso Extremo 1Yesenia Jacqueline Aguilar TarazonaAún no hay calificaciones
- Dos Poemas Del CrisantemoDocumento5 páginasDos Poemas Del CrisantemoUn Bicho Medio RaroAún no hay calificaciones
- Examen Primer Parcial Biologia IDocumento3 páginasExamen Primer Parcial Biologia ISamueel OrthiizAún no hay calificaciones
- Guía de Actividades y Rúbrica de Evaluación - Tarea 1 ReconocimientoDocumento3 páginasGuía de Actividades y Rúbrica de Evaluación - Tarea 1 ReconocimientoDiego Fernando Clavijo PatiñoAún no hay calificaciones
- Los Transgénicos en La MiraDocumento1 páginaLos Transgénicos en La Mirainesvictoria100% (2)
- Como Se Comunican Las Neuronas PDFDocumento43 páginasComo Se Comunican Las Neuronas PDFmiguel50% (2)
- Salida de Campo MalaDocumento33 páginasSalida de Campo MalaAntonio Gómez0% (1)
- ENDYMEDDocumento54 páginasENDYMEDAlexandraAún no hay calificaciones
- Freud PNI de Totem y TabooDocumento1 páginaFreud PNI de Totem y TabooDavidAún no hay calificaciones
- Cuadro Comparativo Neonatos PDFDocumento17 páginasCuadro Comparativo Neonatos PDFLisbeth PalominoAún no hay calificaciones
- Para Que Sirve La OxalaminaDocumento2 páginasPara Que Sirve La OxalaminaSammy FernándezAún no hay calificaciones