También podría gustarte
- Notas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesDe EverandNotas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesAún no hay calificaciones
- TesidDocumento3 páginasTesidIsrael Benoni VallejoAún no hay calificaciones
- Apuntes Tema 4.1Documento14 páginasApuntes Tema 4.1Lucia Menendez RodriguezAún no hay calificaciones
- Sintesis de MelaninaDocumento10 páginasSintesis de MelaninaJesus Octavio Sepulveda CamachoAún no hay calificaciones
- Plantas C4 BNDocumento39 páginasPlantas C4 BNangelAún no hay calificaciones
- Ejemplos de RibozimasDocumento3 páginasEjemplos de RibozimasStefy RecaldeAún no hay calificaciones
- Invaginaciones de La Membrana (Mesosomas y Cromatoforos)Documento37 páginasInvaginaciones de La Membrana (Mesosomas y Cromatoforos)Ingrid karyme fernandez quirozAún no hay calificaciones
- Medicion Del Potencial Hidrico Atraves DDocumento8 páginasMedicion Del Potencial Hidrico Atraves DINGENIOS PANEVIEROS DEL PERU S.A.CAún no hay calificaciones
- Aislamiento de RhizobiumDocumento4 páginasAislamiento de RhizobiumIris Margoth MAún no hay calificaciones
- Proyecto Investigacion Genetica MolecularDocumento9 páginasProyecto Investigacion Genetica MolecularMiguel GonzálesAún no hay calificaciones
- Kalanchoe PinnataDocumento18 páginasKalanchoe PinnataJesus GonzalezAún no hay calificaciones
- Biologia N9Documento6 páginasBiologia N9JOSSEP JARA MUÑOZAún no hay calificaciones
- Nmicrobiol 2016 48 en EsDocumento6 páginasNmicrobiol 2016 48 en EsLonder Vasquez OliveraAún no hay calificaciones
- Práctica 2 GENÉTICADocumento3 páginasPráctica 2 GENÉTICAmiguel angelAún no hay calificaciones
- Seminario 2 Fundamentos de La BMDocumento5 páginasSeminario 2 Fundamentos de La BMAdhari Arango HuamanAún no hay calificaciones
- Sánchez Ramírez Elizabeth Natalí - Guia de Lectura-1Documento5 páginasSánchez Ramírez Elizabeth Natalí - Guia de Lectura-1Elizabeth Natali Sanchez RamirezAún no hay calificaciones
- Código de Barras Del DNA en Cucurbita PepoDocumento27 páginasCódigo de Barras Del DNA en Cucurbita PepoRancel TornezAún no hay calificaciones
- Corrección de ARNDocumento3 páginasCorrección de ARNPaula De la RuaAún no hay calificaciones
- RIBOZIMASDocumento7 páginasRIBOZIMASDaniela OlguinAún no hay calificaciones
- Taller 1 y 2 de BiotecnologíaDocumento4 páginasTaller 1 y 2 de BiotecnologíaR. Daniel RodríguezAún no hay calificaciones
- Texto PresentaciónDocumento7 páginasTexto PresentaciónmariaAún no hay calificaciones
- Bitacora 3Documento7 páginasBitacora 3melani canoAún no hay calificaciones
- Plantas C3 - C4 Grupo 2.en - EsDocumento10 páginasPlantas C3 - C4 Grupo 2.en - EsDavid SantiagoAún no hay calificaciones
- Genética - Formularios de GoogleDocumento16 páginasGenética - Formularios de GoogleLedover Ledover LedoverAún no hay calificaciones
- PIGMENTOS Y MERISTEMOS APICALES DE LAS PLANTAS PRÁCTICA No. 1Documento5 páginasPIGMENTOS Y MERISTEMOS APICALES DE LAS PLANTAS PRÁCTICA No. 1Yosselin NavarroAún no hay calificaciones
- Plástidos DesencadenadosDocumento8 páginasPlástidos DesencadenadosJohana CaezAún no hay calificaciones
- Ilovepdf Merged-2 CompressedDocumento93 páginasIlovepdf Merged-2 CompressedDaniela HazAún no hay calificaciones
- Examen 1Documento14 páginasExamen 1maipo950904Aún no hay calificaciones
- Guía Virtual N°1 Sintesis de Proteinas Del ADN Y ARNDocumento3 páginasGuía Virtual N°1 Sintesis de Proteinas Del ADN Y ARNemilia.nino65Aún no hay calificaciones
- Fenoma VS GenomaDocumento5 páginasFenoma VS GenomaCecilia SanchezAún no hay calificaciones
- Separata de BotanicaDocumento59 páginasSeparata de BotanicaVera Cieza EynerAún no hay calificaciones
- La Rioja - Biologia - 2021Documento14 páginasLa Rioja - Biologia - 2021Paula Gonzalez MesaAún no hay calificaciones
- Plantas c3 y c4Documento12 páginasPlantas c3 y c4carlosAún no hay calificaciones
- CloroplastosDocumento3 páginasCloroplastosJoss AndradeAún no hay calificaciones
- Tema 4Documento25 páginasTema 4EDISONAún no hay calificaciones
- Bioquímica Vegetal CuestionarioDocumento17 páginasBioquímica Vegetal CuestionarioKatherine GaitánAún no hay calificaciones
- Práctica 13 Laboratorio BioquímicaDocumento9 páginasPráctica 13 Laboratorio BioquímicaJhon AcerosAún no hay calificaciones
- Monografía BriofitasDocumento11 páginasMonografía Briofitaschidote563Aún no hay calificaciones
- ProtoplastoDocumento7 páginasProtoplastoFabricio JácomeAún no hay calificaciones
- EL ARN y Su ImportanciaDocumento10 páginasEL ARN y Su ImportanciaDarwinAún no hay calificaciones
- Origen Evol y Tax y Citoplasma Bacteriano 2021Documento40 páginasOrigen Evol y Tax y Citoplasma Bacteriano 2021Victor Reyes FuenzalidaAún no hay calificaciones
- La Rizobacteria Promotora Del Crecimiento Vegetal Arthrobacter Agilis Umcv2 Coloniza Endofíticamente A Medicago TruncatulaDocumento6 páginasLa Rizobacteria Promotora Del Crecimiento Vegetal Arthrobacter Agilis Umcv2 Coloniza Endofíticamente A Medicago TruncatulaaironixxAún no hay calificaciones
- Cianobacterias PDFDocumento16 páginasCianobacterias PDFEurogunzReyes100% (1)
- Qué Desventaja Tiene Poner Un Campo Más Con Las Descripciones de Los Términos de Gene Ontology en Esta TablaDocumento4 páginasQué Desventaja Tiene Poner Un Campo Más Con Las Descripciones de Los Términos de Gene Ontology en Esta TablaDavid AvilésAún no hay calificaciones
- Actividad Evaluativa 6Documento7 páginasActividad Evaluativa 6Kevin MalagonAún no hay calificaciones
- 16 Tarea Bioqupimica 22Documento5 páginas16 Tarea Bioqupimica 22Yazmin MartinezAún no hay calificaciones
- TALLER DE BIOLOGIA - IcfesDocumento4 páginasTALLER DE BIOLOGIA - IcfesVICTOR PEÑAAún no hay calificaciones
- Trabajo Biologia Modulo I Lechuga EQUIPO 5Documento8 páginasTrabajo Biologia Modulo I Lechuga EQUIPO 5maria reyesAún no hay calificaciones
- Genética Molecular AlumnosDocumento48 páginasGenética Molecular AlumnosNydheAún no hay calificaciones
- Investigacion TerpenosDocumento8 páginasInvestigacion TerpenosKatheryne SimbañaAún no hay calificaciones
- Anatomia FoliarDocumento4 páginasAnatomia FoliarAngelica Margarito CanoAún no hay calificaciones
- BacteriorrodopsinaDocumento11 páginasBacteriorrodopsinaLizitha Mora SotoAún no hay calificaciones
- Semana 8 - Ficha 2Documento4 páginasSemana 8 - Ficha 2Sofia Raquel Abanto A.Aún no hay calificaciones
- Ensayo de Plantas c3 c4 y CAM Original TaniaDocumento7 páginasEnsayo de Plantas c3 c4 y CAM Original Taniadariela garcia0% (2)
- Genoma BacterianoDocumento7 páginasGenoma BacterianoSaul Adrian Mamani SanchezAún no hay calificaciones
- Texto Biologia Molecular 2012Documento81 páginasTexto Biologia Molecular 2012Estela JerezAún no hay calificaciones
- Sistema de Dos SuperreinosDocumento5 páginasSistema de Dos SuperreinosEskereAún no hay calificaciones
- QF - PRÁCTICA 13 LABORATORIO BIOQUÍMICA - Acidos Nucleicos - Aceros, Gutierrez, Martinez - Grupo 3Documento8 páginasQF - PRÁCTICA 13 LABORATORIO BIOQUÍMICA - Acidos Nucleicos - Aceros, Gutierrez, Martinez - Grupo 3Jhon AcerosAún no hay calificaciones
- Guia 3 Biologia 9 PDFDocumento7 páginasGuia 3 Biologia 9 PDFeymi diazAún no hay calificaciones
- Caracterizacion Molecular de Papas Nativas en Ayacucho Mediante AFLPDocumento6 páginasCaracterizacion Molecular de Papas Nativas en Ayacucho Mediante AFLPYOEL IDROGO HUAMANAún no hay calificaciones
- TRIPTICO SpallanzaniDocumento2 páginasTRIPTICO SpallanzaniIsrael Benoni VallejoAún no hay calificaciones
- TripticoDocumento2 páginasTripticoIsrael Benoni VallejoAún no hay calificaciones
- PRÁCTICA 1 Identificacion de Bioelementos IsraelDocumento7 páginasPRÁCTICA 1 Identificacion de Bioelementos IsraelIsrael Benoni Vallejo100% (1)
- Frankenberg - Teoria-CriticaDocumento18 páginasFrankenberg - Teoria-CriticaMario VelaAún no hay calificaciones
- Material de VidrioDocumento7 páginasMaterial de VidrioIsrael Benoni VallejoAún no hay calificaciones
- 2014 Naturales y de SaludDocumento12 páginas2014 Naturales y de SaludIsrael Benoni VallejoAún no hay calificaciones
- Copia de Relacion de La Quimica Con La TecnologiaDocumento1 páginaCopia de Relacion de La Quimica Con La TecnologiaIsrael Benoni VallejoAún no hay calificaciones
- Dibujo - Técnicas - Cómo Dibujar Manga - Libro 1 - Personajes - 110 PagDocumento110 páginasDibujo - Técnicas - Cómo Dibujar Manga - Libro 1 - Personajes - 110 PagPercy Castro V.93% (92)
- Practicar Sync LinuxDocumento3 páginasPracticar Sync Linuxkabuto cAún no hay calificaciones
- Cancion Al Arbol Del OlvidoDocumento3 páginasCancion Al Arbol Del OlvidoMirii Kolakau50% (4)
- Descubre y Entiende de Una Vez La Estrecha Relación Entre Cloro y PH en PiscinasDocumento9 páginasDescubre y Entiende de Una Vez La Estrecha Relación Entre Cloro y PH en PiscinasJuan GarciaAún no hay calificaciones
- S4.Estructura Cristalina - Defectos Estructurales 20-1Documento14 páginasS4.Estructura Cristalina - Defectos Estructurales 20-1Paolo Chavez CernaAún no hay calificaciones
- Pies en Polvorosa ComentarioDocumento9 páginasPies en Polvorosa ComentariocepermotrilAún no hay calificaciones
- El Ciclo de Krebs, Vía Central Del Metabolismo de Los CarbohidratosDocumento18 páginasEl Ciclo de Krebs, Vía Central Del Metabolismo de Los CarbohidratosJUanAún no hay calificaciones
- Estructuras de ConcretoDocumento27 páginasEstructuras de ConcretoTadeo SanchezAún no hay calificaciones
- Proyecto FinalDocumento37 páginasProyecto FinalTania RamírezAún no hay calificaciones
- Normas Específicas de Bioseguridad para Desechos HospitalariosDocumento8 páginasNormas Específicas de Bioseguridad para Desechos HospitalariosMario PérezAún no hay calificaciones
- Métodos y Modelos para La Predicción y Selección Genómica en El Mejoramiento de PlantasDocumento2 páginasMétodos y Modelos para La Predicción y Selección Genómica en El Mejoramiento de Plantasp3dr0javaAún no hay calificaciones
- Reproduccion AsexualDocumento2 páginasReproduccion AsexualFernandaAún no hay calificaciones
- COMUNICACIONDocumento13 páginasCOMUNICACIONFreyssi Zelada AlcaldeAún no hay calificaciones
- 6 Año - Historia - Guía - Ambientes Naturales en ChileDocumento4 páginas6 Año - Historia - Guía - Ambientes Naturales en ChileMelania Varela ZabalaAún no hay calificaciones
- Hoja de Trabajo 2da. Evaluac...Documento10 páginasHoja de Trabajo 2da. Evaluac...Jennifer PresencinAún no hay calificaciones
- Menús Tradición 2024Documento21 páginasMenús Tradición 2024Racam10Aún no hay calificaciones
- Proforma Tropipalma SaDocumento3 páginasProforma Tropipalma SaJhoz Alexander Cantos LitumaAún no hay calificaciones
- Simulacro AeropuertoDocumento3 páginasSimulacro AeropuertoFederico RolandiAún no hay calificaciones
- Informe Gerente PickingDocumento13 páginasInforme Gerente PickingClaudia BgAún no hay calificaciones
- Resumen de Numeros Enteros Paes 2023Documento11 páginasResumen de Numeros Enteros Paes 2023Manuel AlarconAún no hay calificaciones
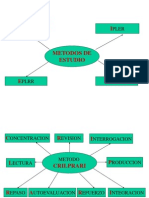
- Metodos de EstudioDocumento23 páginasMetodos de Estudiorichtr100% (1)
- Embrague Multidisco (Videos Embrague y Controles de Embrague)Documento11 páginasEmbrague Multidisco (Videos Embrague y Controles de Embrague)Emanuel SanchezAún no hay calificaciones
- Distitron 5119 Elsxq F.T. - EspDocumento1 páginaDistitron 5119 Elsxq F.T. - Espsara.mebautoAún no hay calificaciones
- Hemorragia DigestivaDocumento59 páginasHemorragia Digestivatitolivio89Aún no hay calificaciones
- Calculo de RMC (Método Gráfico)Documento1 páginaCalculo de RMC (Método Gráfico)panda.gamer.yt402Aún no hay calificaciones
- Trifolio Quimica 3Documento3 páginasTrifolio Quimica 3Dariana PadillaAún no hay calificaciones
- Reseña Critica "Volver Al Futuro"Documento2 páginasReseña Critica "Volver Al Futuro"Darkzone0% (2)
- EducamosCLM Seguimiento EducativoDocumento1 páginaEducamosCLM Seguimiento EducativoJuan José Bernal ParraAún no hay calificaciones
- Niversidad Tecnologica Regional Del Sur: Asignatura: Administración de La ProducciónDocumento3 páginasNiversidad Tecnologica Regional Del Sur: Asignatura: Administración de La Producciónestefani dzulAún no hay calificaciones
- Transmision Señales ConceptosDocumento10 páginasTransmision Señales Conceptosmetaphysics18Aún no hay calificaciones
- Nop Lasbambas 20 02 Dosaje EtilicoDocumento6 páginasNop Lasbambas 20 02 Dosaje EtilicofrancisAún no hay calificaciones
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- Homo antecessor: El nacimiento de una especieDe EverandHomo antecessor: El nacimiento de una especieCalificación: 5 de 5 estrellas5/5 (1)
- GuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisDe EverandGuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisCalificación: 4 de 5 estrellas4/5 (9)
- 50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaDe Everand50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaCalificación: 4 de 5 estrellas4/5 (37)
- Notas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesDe EverandNotas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesAún no hay calificaciones
- Neuropsicología: Los fundamentos de la materiaDe EverandNeuropsicología: Los fundamentos de la materiaCalificación: 5 de 5 estrellas5/5 (1)
- El cerebro del niño explicado a los padresDe EverandEl cerebro del niño explicado a los padresCalificación: 4.5 de 5 estrellas4.5/5 (147)
- Nuestra mente nos engaña: Sesgos y errores cognitivos que todos cometemosDe EverandNuestra mente nos engaña: Sesgos y errores cognitivos que todos cometemosCalificación: 3.5 de 5 estrellas3.5/5 (6)
- Sacale partido a tu cerebro: Todo lo que necesitas saber para mejorar tu memoria, tomar decisiones y aprovechar todo tu potencialDe EverandSacale partido a tu cerebro: Todo lo que necesitas saber para mejorar tu memoria, tomar decisiones y aprovechar todo tu potencialCalificación: 3 de 5 estrellas3/5 (1)
- Minerales y vitaminas: Micronutrientes esenciales en la alimentación, nutrición y salud.De EverandMinerales y vitaminas: Micronutrientes esenciales en la alimentación, nutrición y salud.Calificación: 3 de 5 estrellas3/5 (2)
- Inteligencia artificial: Una exploración filosófica sobre el futuro de la mente y la concienciaDe EverandInteligencia artificial: Una exploración filosófica sobre el futuro de la mente y la concienciaAna Isabel Sánchez DíezCalificación: 4 de 5 estrellas4/5 (3)
- Los cinco ritos tibetanos de la eterna juventud: El ojo de la revelaciónDe EverandLos cinco ritos tibetanos de la eterna juventud: El ojo de la revelaciónCalificación: 4.5 de 5 estrellas4.5/5 (12)
- Curso Práctico de Química GeneralDe EverandCurso Práctico de Química GeneralCalificación: 4 de 5 estrellas4/5 (4)
- Cerebros rotos: Pacientes asombrosos que me enseñaron a vivirDe EverandCerebros rotos: Pacientes asombrosos que me enseñaron a vivirCalificación: 5 de 5 estrellas5/5 (3)
- 200 tareas en terapia breve: 2ª ediciónDe Everand200 tareas en terapia breve: 2ª ediciónCalificación: 4.5 de 5 estrellas4.5/5 (33)
- Abrázame fuerte: Siete conversaciones para un amor duraderoDe EverandAbrázame fuerte: Siete conversaciones para un amor duraderoManu BerásteguiCalificación: 4.5 de 5 estrellas4.5/5 (13)
- Cómo dormir bien: Cómo reconocer, prevenir y tratar las dificultades para dormir bienDe EverandCómo dormir bien: Cómo reconocer, prevenir y tratar las dificultades para dormir bienCalificación: 5 de 5 estrellas5/5 (1)
- El perfil psicológico de Jesús: Aprendamos del Maestro a manejar efectivamente nuestras emocionesDe EverandEl perfil psicológico de Jesús: Aprendamos del Maestro a manejar efectivamente nuestras emocionesCalificación: 4 de 5 estrellas4/5 (4)
- Sesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesDe EverandSesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesCalificación: 4.5 de 5 estrellas4.5/5 (13)
- Flujo de fluidos e intercambio de calorDe EverandFlujo de fluidos e intercambio de calorCalificación: 1 de 5 estrellas1/5 (1)
- El autismo: Reflexiones y pautas para comprenderlo y abordarloDe EverandEl autismo: Reflexiones y pautas para comprenderlo y abordarloCalificación: 4 de 5 estrellas4/5 (7)
- Fisicoquímica II: Materia, electricidad y magnetismo. Fuerzas y camposDe EverandFisicoquímica II: Materia, electricidad y magnetismo. Fuerzas y camposAún no hay calificaciones