También podría gustarte
- Articulo de Nucleo CelularDocumento13 páginasArticulo de Nucleo CelularMishelle HuamaníAún no hay calificaciones
- Transporte Entre El Núcleo y El Citoplasma.Documento3 páginasTransporte Entre El Núcleo y El Citoplasma.Alejandra SánchezAún no hay calificaciones
- Lodish - Capitulo16Documento52 páginasLodish - Capitulo16t1020400721Aún no hay calificaciones
- Reguladores nuclearesDocumento5 páginasReguladores nuclearesLorena Castillo ValtierraAún no hay calificaciones
- Control expresión génica celularDocumento7 páginasControl expresión génica celularTony MartínezAún no hay calificaciones
- 2do Parcial Medicina MolecularDocumento7 páginas2do Parcial Medicina MolecularAna Daniela Cruz ZarabiaAún no hay calificaciones
- Transcripción Del DNA en Procariotas y Su RegulaciónDocumento30 páginasTranscripción Del DNA en Procariotas y Su RegulaciónRaquel Pérez FernándezAún no hay calificaciones
- Membrana NuclearDocumento54 páginasMembrana NuclearLuis David Bello RodríguezAún no hay calificaciones
- Sindrome de GordonDocumento9 páginasSindrome de Gordonangelica39Aún no hay calificaciones
- Informe S9Documento5 páginasInforme S9Valeria DelpinoAún no hay calificaciones
- Mecanismos genómicos y no genómicos de los corticoidesDocumento4 páginasMecanismos genómicos y no genómicos de los corticoidesAnonymous UQkAWZ100% (1)
- AldosteronaDocumento13 páginasAldosteronaAzu G CenalAún no hay calificaciones
- Receptores NuclearesDocumento5 páginasReceptores NuclearescindyAún no hay calificaciones
- BioquimicaDocumento13 páginasBioquimicaCome GalletasAún no hay calificaciones
- Resumen Biologia MolecularDocumento12 páginasResumen Biologia MolecularMaria Gabriela Urdaneta ZubalevichAún no hay calificaciones
- GeneticaDocumento48 páginasGeneticafernanda lemusAún no hay calificaciones
- Ciclo Celular Genes Checkpoint y CáncerDocumento19 páginasCiclo Celular Genes Checkpoint y CáncerEdgar A. DominguezAún no hay calificaciones
- Regulación transcripción genes nutriciónDocumento5 páginasRegulación transcripción genes nutriciónDiana Elizabeth Hernandez FloresAún no hay calificaciones
- Epigenetica ArtDocumento6 páginasEpigenetica ArtJJ James VPAún no hay calificaciones
- Receptor Tirosina QuinasaDocumento10 páginasReceptor Tirosina QuinasaSebastian MartinezAún no hay calificaciones
- ESTRUCTURA Y FUNCIONES DEL NUCLEO CELULARDocumento36 páginasESTRUCTURA Y FUNCIONES DEL NUCLEO CELULARsabrina100% (1)
- Epigenética AndrésDocumento30 páginasEpigenética AndrésAlberto BeskowAún no hay calificaciones
- Funciones Biológicas de Los Ácidos NucleicosDocumento4 páginasFunciones Biológicas de Los Ácidos NucleicosMariana crystel Cabanillas SalcidoAún no hay calificaciones
- Funciones NucleoDocumento5 páginasFunciones NucleoluisAún no hay calificaciones
- Núcleo CelularDocumento42 páginasNúcleo CelularmarioAún no hay calificaciones
- Biologia Molecular. Fundamentos y AplicacionesDocumento173 páginasBiologia Molecular. Fundamentos y Aplicacionesshady182100% (1)
- Info ExpoDocumento4 páginasInfo ExpoMaria VadilloAún no hay calificaciones
- Regulación Génica en EucariotesDocumento44 páginasRegulación Génica en EucariotesMauricio SanchezAún no hay calificaciones
- Regulación de GenesDocumento51 páginasRegulación de GenesFermín Armando Ledezma VelázquezAún no hay calificaciones
- Activación y Represión Transcripcional, Elementos: - A-Vistos en PrácticasDocumento57 páginasActivación y Represión Transcripcional, Elementos: - A-Vistos en PrácticasmimotronicAún no hay calificaciones
- LN Cap 1.6Documento40 páginasLN Cap 1.6Daniela Patricia Alvarez AravenaAún no hay calificaciones
- Senalizacion IntercelularDocumento35 páginasSenalizacion IntercelularIrving ParraAún no hay calificaciones
- Capítulo 1Documento21 páginasCapítulo 1John DittelAún no hay calificaciones
- Farmacologia MolecularDocumento5 páginasFarmacologia MolecularolgakatherineAún no hay calificaciones
- Regulación de La Expresión Génica EucariotasDocumento20 páginasRegulación de La Expresión Génica EucariotasGloribel Bolívar100% (1)
- Organización y Regulación de La Transcripción GenéticaDocumento7 páginasOrganización y Regulación de La Transcripción GenéticaorutraodnamraAún no hay calificaciones
- Biologia SEMANA 15Documento6 páginasBiologia SEMANA 15john salchichon100% (6)
- Capitulo 1. Embriología MédicaDocumento16 páginasCapitulo 1. Embriología MédicaYessenia MillaAún no hay calificaciones
- TCR 1Documento14 páginasTCR 1Kevin alberto Pérez de la cruzAún no hay calificaciones
- Guía de Estudio para El Cuarto Parcial de Biología MolecularDocumento9 páginasGuía de Estudio para El Cuarto Parcial de Biología MolecularMolina RafaelAún no hay calificaciones
- Nutrigenómica y nutrigenética: la base genética de la nutriciónDocumento53 páginasNutrigenómica y nutrigenética: la base genética de la nutriciónMario MoralesAún no hay calificaciones
- Las Moleculas de Adhesion y La Respuesta Inmune PDFDocumento8 páginasLas Moleculas de Adhesion y La Respuesta Inmune PDFAlexa BuenoAún no hay calificaciones
- Regulación de La Expresión GénicaDocumento34 páginasRegulación de La Expresión GénicamiriammoreyAún no hay calificaciones
- Telomeros y TelomerasasDocumento4 páginasTelomeros y TelomerasasLuz Vanessa ParejoAún no hay calificaciones
- Farmacología de los receptores nuclearesDocumento21 páginasFarmacología de los receptores nuclearesMagnolia VelizAún no hay calificaciones
- Regulación génica pre-transcripcionalDocumento10 páginasRegulación génica pre-transcripcionaltaricura estebanAún no hay calificaciones
- Tirosina QuinasaDocumento6 páginasTirosina QuinasaSheyla Jara TorresAún no hay calificaciones
- Semana 4 PDFDocumento4 páginasSemana 4 PDFMiriam González RuizAún no hay calificaciones
- Aspectos Bioquimicos de La Acción HormonalDocumento12 páginasAspectos Bioquimicos de La Acción HormonalJosue Alvites Ramos100% (1)
- Introducción a la genómica grupo 01Documento10 páginasIntroducción a la genómica grupo 01Armando MoraAún no hay calificaciones
- Bioquimica - Receptores de MembranaDocumento6 páginasBioquimica - Receptores de MembranaGeison MartinsAún no hay calificaciones
- Transcripción y Traducción en Organismos ProcariotasDocumento2 páginasTranscripción y Traducción en Organismos ProcariotasLuis Mario MartinezAún no hay calificaciones
- Trabajo EpigenómicaDocumento12 páginasTrabajo EpigenómicapepaAún no hay calificaciones
- 5.expresion Genetica y NeurotransmisoresDocumento43 páginas5.expresion Genetica y NeurotransmisoresJoshein PerezAún no hay calificaciones
- Taller Sobre Lectura Crítica - Ana Milena RuizDocumento4 páginasTaller Sobre Lectura Crítica - Ana Milena RuizAna Milena Ruiz RomeroAún no hay calificaciones
- Regulación transcripcional eucariota: activación, remodelado cromatínico y modificaciones histonasDocumento34 páginasRegulación transcripcional eucariota: activación, remodelado cromatínico y modificaciones histonasRaquel Pérez FernándezAún no hay calificaciones
- Alteraciones en La Vía MAPKDocumento9 páginasAlteraciones en La Vía MAPKVíctor Bravo PAún no hay calificaciones
- Receptores EndocrinosDocumento5 páginasReceptores EndocrinosNayely ErasAún no hay calificaciones
- Más allá del ADN: La Revolución Epigenética: Desde Mecanismos Celulares hasta Factores Ambientales: Cómo la Epigenética Moldea Nuestro Destino Biológico y las Implicaciones para la Salud, el Comportamiento y el Futuro de la InvestigaciónDe EverandMás allá del ADN: La Revolución Epigenética: Desde Mecanismos Celulares hasta Factores Ambientales: Cómo la Epigenética Moldea Nuestro Destino Biológico y las Implicaciones para la Salud, el Comportamiento y el Futuro de la InvestigaciónAún no hay calificaciones
- INFO - Potencial de Membrana CelularDocumento36 páginasINFO - Potencial de Membrana CelularOscar Andres Diaz GonzalezAún no hay calificaciones
- Manual Biología CelularDocumento92 páginasManual Biología Celularjose cardenasAún no hay calificaciones
- INFO Cadena de Transporte de Electrones y FosforilaciónDocumento9 páginasINFO Cadena de Transporte de Electrones y FosforilaciónOscar Andres Diaz GonzalezAún no hay calificaciones
- Transporte VesicularDocumento9 páginasTransporte VesicularkerenAún no hay calificaciones
- Clase 2Documento13 páginasClase 2Francisco Bladimir Ramírez BeltránAún no hay calificaciones
- Mitos y verdades del ejercicioDocumento17 páginasMitos y verdades del ejercicioffAún no hay calificaciones
- La Inmortalidad Procaritica y La Tenacidad de La VidaDocumento8 páginasLa Inmortalidad Procaritica y La Tenacidad de La VidaJenifer AcevesAún no hay calificaciones
- Lesión CélularDocumento9 páginasLesión CélularkerenAún no hay calificaciones
- Capítulo 8.2 Suplementos de ProteínaDocumento34 páginasCapítulo 8.2 Suplementos de Proteínajesuscanocano100% (1)
- Compuesto en El Cuerpo HumanoDocumento4 páginasCompuesto en El Cuerpo HumanoMarvin OrtizAún no hay calificaciones
- Adan Biblico y Cientifico - Un Vern S. PoythressDocumento79 páginasAdan Biblico y Cientifico - Un Vern S. PoythressEmmanuel Gonzalez ReyesAún no hay calificaciones
- Quinoa Case Study EsDocumento86 páginasQuinoa Case Study EsNohemy KpaAún no hay calificaciones
- Distinguir La Ficción La Verdad Sobre El Zumo de NoniDocumento19 páginasDistinguir La Ficción La Verdad Sobre El Zumo de NoniLuisa Fernanda Herrera AgudeloAún no hay calificaciones
- Práctica #5Documento6 páginasPráctica #5Celeste Ramos ChavarryAún no hay calificaciones
- Obtención de Cuero Box Calf Al Cromo de Piel Adulta de OvejaDocumento17 páginasObtención de Cuero Box Calf Al Cromo de Piel Adulta de OvejaAdela Huarahuara CruzAún no hay calificaciones
- CLICAP 2005 - Memorias - Resúmenes de InvestigaciónDocumento213 páginasCLICAP 2005 - Memorias - Resúmenes de InvestigaciónrvergneAún no hay calificaciones
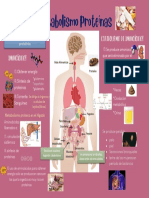
- Metabolismo ProteínasDocumento1 páginaMetabolismo Proteínassokodas hijo de sukedas57% (7)
- RESUMEN de CLASES DE BIOLOGÍA CelularDocumento23 páginasRESUMEN de CLASES DE BIOLOGÍA CelularKenya San JuanAún no hay calificaciones
- 2 Contexto Internacional 2Documento42 páginas2 Contexto Internacional 2Jhon WillAún no hay calificaciones
- Practica 3 Biologia MolecularDocumento11 páginasPractica 3 Biologia MolecularQuetzally ValdiviaAún no hay calificaciones
- Protocolos HemodialisisDocumento94 páginasProtocolos HemodialisisantonioAún no hay calificaciones
- Dirks2015 en EsDocumento10 páginasDirks2015 en EsDaniel IturriagaAún no hay calificaciones
- Preguntas Tipo Prueba ArqueometríaDocumento21 páginasPreguntas Tipo Prueba ArqueometríaMAGDALENA MARDONESAún no hay calificaciones
- Cuerpo EndomorfoDocumento88 páginasCuerpo EndomorfoDanisa Gaitan100% (1)
- Diseñan Una Proteína Artificial Capaz de Filtrar y Degradar PlásticosDocumento2 páginasDiseñan Una Proteína Artificial Capaz de Filtrar y Degradar PlásticosMaria Jose NicolaldeAún no hay calificaciones
- Extraccion de Material Genetico en Spinacia OleraceaDocumento5 páginasExtraccion de Material Genetico en Spinacia Oleraceaxnew_67026911Aún no hay calificaciones
- Proyecto QuinuaDocumento98 páginasProyecto QuinuaMrRODRIGORC83% (6)
- Buffer de Los AminoácidosDocumento8 páginasBuffer de Los AminoácidosanrealdoAún no hay calificaciones
- Vademecum Abbot PDFDocumento72 páginasVademecum Abbot PDFLina María Giraldo Ospina33% (3)
- Tratamiento nutricional propuesto para pacientes hepáticosDocumento98 páginasTratamiento nutricional propuesto para pacientes hepáticosmacario mamaniAún no hay calificaciones
- Cuaderno Del Alumno Nutricion 2021Documento187 páginasCuaderno Del Alumno Nutricion 2021NAGU SAPPAún no hay calificaciones
- Biología: Química de la materia vivaDocumento6 páginasBiología: Química de la materia vivaDenner Kilder Cabrejos RamirezAún no hay calificaciones
- Historia de la ciencia de los alimentosDocumento21 páginasHistoria de la ciencia de los alimentosSandro samanamudAún no hay calificaciones
- Exposicion Teoria CelularDocumento4 páginasExposicion Teoria CelularJUAN ALEX FERNANDO CHITIVA VERGARAAún no hay calificaciones