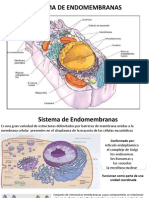
También podría gustarte
- Cap. 12 Celula BiologiaDocumento54 páginasCap. 12 Celula BiologiaEmiliano FrancoAún no hay calificaciones
- Modulo 8-04Documento17 páginasModulo 8-04Gustavo RaeleAún no hay calificaciones
- Sistema de EndomembranasDocumento9 páginasSistema de Endomembranasandrea rochaAún no hay calificaciones
- RibosomasDocumento6 páginasRibosomaspakalna80% (5)
- CLASE 6 Reticulo Endoplasmatico MEDICINA 2021 FINALDocumento23 páginasCLASE 6 Reticulo Endoplasmatico MEDICINA 2021 FINALya tu sabeAún no hay calificaciones
- Copia de Sistemas de EndomembranasDocumento8 páginasCopia de Sistemas de EndomembranasMartina AgueroAún no hay calificaciones
- EucariotasDocumento49 páginasEucariotasMohamed AléAún no hay calificaciones
- Sistema de EndomembranasDocumento4 páginasSistema de EndomembranasJosephGabrielAún no hay calificaciones
- Transporte y Relaciones - Membrana PDFDocumento12 páginasTransporte y Relaciones - Membrana PDFBritney Charline AscherAún no hay calificaciones
- EndomembranaDocumento4 páginasEndomembranaJonatham R JiménezAún no hay calificaciones
- MEMBRANAS INTERNAS 1 - TeoríaDocumento25 páginasMEMBRANAS INTERNAS 1 - TeoríaMarino GuerreroAún no hay calificaciones
- Unid 2-Sistemas-de-Endomembranas-docx - GR 6ºDocumento7 páginasUnid 2-Sistemas-de-Endomembranas-docx - GR 6ºMARITZA CORTES PARRAAún no hay calificaciones
- Organizacion de Las CelulasDocumento9 páginasOrganizacion de Las CelulasEider Daniel Pastuzan DelgadoAún no hay calificaciones
- 24 EndomembranaDocumento4 páginas24 EndomembranaFreddy David HerreraAún no hay calificaciones
- Estructura y Función CelularDocumento4 páginasEstructura y Función CelularCarolina OrtizAún no hay calificaciones
- Resumen de Biologia Celular - Agosto.2021Documento8 páginasResumen de Biologia Celular - Agosto.2021Michell AndradeAún no hay calificaciones
- 3.membrana Citoplasmica Parte 2 BDocumento11 páginas3.membrana Citoplasmica Parte 2 BMonseAún no hay calificaciones
- Célula AnimalDocumento17 páginasCélula AnimalJUDITH ALONDRA TORRES HERNANDEZAún no hay calificaciones
- TEMA II. Sistemas de Membr. Citoplásmica. Resp. Aeróbica y La Mitoco. Citoesqueleto y Mov. Cel.Documento45 páginasTEMA II. Sistemas de Membr. Citoplásmica. Resp. Aeróbica y La Mitoco. Citoesqueleto y Mov. Cel.BELL TOMLINSONAún no hay calificaciones
- La CelulaDocumento4 páginasLa CelulaKarolyne RealAún no hay calificaciones
- Unidad Didáctica IvDocumento10 páginasUnidad Didáctica Ivtrasteandopuntocom tpcAún no hay calificaciones
- Citoplasma CelularDocumento19 páginasCitoplasma CelularAlisson Caicedo100% (2)
- Cuestionario Sistema de EndomembranasDocumento4 páginasCuestionario Sistema de EndomembranasLeonora PaelAún no hay calificaciones
- Tarea V Funciones de Cada Una de Las Estructuras CelularesDocumento2 páginasTarea V Funciones de Cada Una de Las Estructuras CelularesEddy Liranzo FañaAún no hay calificaciones
- Informe EditadoDocumento27 páginasInforme EditadoErick SaldañaAún no hay calificaciones
- Tráfico VesicularDocumento55 páginasTráfico VesicularAficionadoAún no hay calificaciones
- Fisio Primer ParcialDocumento34 páginasFisio Primer ParcialFlor SternariAún no hay calificaciones
- Bic U2 t3 PDF 18Documento5 páginasBic U2 t3 PDF 18bombitopazAún no hay calificaciones
- Script 4ta PresentacionDocumento3 páginasScript 4ta PresentacionLukas Felipe Rey RicardoAún no hay calificaciones
- 8.sistemas Membranosos de La Célula EucariotaDocumento9 páginas8.sistemas Membranosos de La Célula EucariotaAntony VillamarAún no hay calificaciones
- Neurociencias ResumenesDocumento15 páginasNeurociencias ResumenesMelany Aldana DAIYOUBAún no hay calificaciones
- 11-Sistema de Endomembranas-Apunte GeneralDocumento19 páginas11-Sistema de Endomembranas-Apunte GeneralTomas Scialfa BenderskyAún no hay calificaciones
- Células Eucariotas y ProcariotasDocumento5 páginasCélulas Eucariotas y ProcariotasFanny CMunvAún no hay calificaciones
- Guia de HistologiaDocumento12 páginasGuia de Histologiayexiimar16Aún no hay calificaciones
- Celula AnimaDocumento5 páginasCelula Animapo poAún no hay calificaciones
- La CelulaDocumento6 páginasLa CelulaYusiley Zeballos MejiaAún no hay calificaciones
- Organelos CelularesDocumento38 páginasOrganelos Celularesyoselin gonzalezAún no hay calificaciones
- Biología - Química UNED, Tema4Documento6 páginasBiología - Química UNED, Tema4Marta Arias FernandezAún no hay calificaciones
- MitocondriasDocumento6 páginasMitocondriasemmanuelsojuelsilvaAún no hay calificaciones
- Guía de Organelos CelularesDocumento8 páginasGuía de Organelos CelularesCamila Bossio CastillaAún no hay calificaciones
- Modulo III Reticulo Endoplásmicosistema de Endomembranas Con Audio Final 2023Documento29 páginasModulo III Reticulo Endoplásmicosistema de Endomembranas Con Audio Final 2023Madelin BolañosAún no hay calificaciones
- La Membrana PDFDocumento31 páginasLa Membrana PDFCopycentro y Equipos MaryAún no hay calificaciones
- Tema 3 Estructura y Función CelularDocumento8 páginasTema 3 Estructura y Función CelularJorge GarcíaAún no hay calificaciones
- Notas BiologiaDocumento3 páginasNotas BiologiaDayana VasquezAún no hay calificaciones
- SVC Biología 08Documento7 páginasSVC Biología 08Ptricia OliverosAún no hay calificaciones
- Organelos CitoplasmáticosDocumento24 páginasOrganelos CitoplasmáticosAlbaniBriceñoAún no hay calificaciones
- 03 Premedicina Biologia+Celular+y+Molecular Celula-Organelos+Celulares-transporte+MembranalDocumento9 páginas03 Premedicina Biologia+Celular+y+Molecular Celula-Organelos+Celulares-transporte+Membranalgeorgepiste1Aún no hay calificaciones
- Taller de BiologiaDocumento10 páginasTaller de BiologiaYuveidys Chamorro VergaraAún no hay calificaciones
- Las Entidades Fundamentales de Todo Ser Vivo Son Las CélulasDocumento5 páginasLas Entidades Fundamentales de Todo Ser Vivo Son Las CélulasroximarcontrerasAún no hay calificaciones
- BiologiaDocumento156 páginasBiologiaKatherine AltamiranoAún no hay calificaciones
- CELULADocumento108 páginasCELULAalejandraAún no hay calificaciones
- Biología Unidad 5-Estructura y Función CelularDocumento11 páginasBiología Unidad 5-Estructura y Función CelularJenny MoreiraAún no hay calificaciones
- Apuntes Estructura CelularDocumento12 páginasApuntes Estructura CelularMaria Teresa MolinaAún no hay calificaciones
- Capitulo 12 AlbertsDocumento35 páginasCapitulo 12 AlbertsRicardo Cuatlayotl OlarteAún no hay calificaciones
- Componentes de Endom 184417 Downloadable 5486838Documento11 páginasComponentes de Endom 184417 Downloadable 5486838evelynpapadopulos1Aún no hay calificaciones
- Separata Organelas y Componentes Extracelulares de Una Celula EucariotaDocumento9 páginasSeparata Organelas y Componentes Extracelulares de Una Celula Eucariotalucero liraAún no hay calificaciones
- Los Ribosomas - BiologiaDocumento7 páginasLos Ribosomas - BiologiaMag SosaAún no hay calificaciones
- Anatomía y Fisiología CelularDocumento7 páginasAnatomía y Fisiología CelularDALVIC ALARCON50% (2)
- Unidad 2b Organelos y Trasporte EndomembranalDocumento19 páginasUnidad 2b Organelos y Trasporte EndomembranalArgeliaAún no hay calificaciones
- Taller Informacion en InternetDocumento9 páginasTaller Informacion en InternetDIBL 96Aún no hay calificaciones
- Ciclo Celular: La Interfase Se Divide en Etapas: G G1, S y G2. Desde El Punto de Vista de La GenéticaDocumento11 páginasCiclo Celular: La Interfase Se Divide en Etapas: G G1, S y G2. Desde El Punto de Vista de La GenéticaDIBL 96Aún no hay calificaciones
- Guia Observación Celular BMS1100Documento10 páginasGuia Observación Celular BMS1100DIBL 96Aún no hay calificaciones
- Trabajo QuimicaDocumento4 páginasTrabajo QuimicaDIBL 96Aún no hay calificaciones
- Guía Huesos Y Sus Funciones: Escuela de SaludDocumento19 páginasGuía Huesos Y Sus Funciones: Escuela de SaludDIBL 96Aún no hay calificaciones
- Trabajo InfoDocumento6 páginasTrabajo InfoDIBL 96Aún no hay calificaciones
- GLANDULAS Y HORMONAS (A)Documento10 páginasGLANDULAS Y HORMONAS (A)DIBL 96Aún no hay calificaciones
- Estudio de CompetenciaDocumento1 páginaEstudio de CompetenciaDIBL 96Aún no hay calificaciones
- Oportunidades e Ideas 102Documento6 páginasOportunidades e Ideas 102DIBL 96Aún no hay calificaciones
- DIGESTIVO AlDocumento10 páginasDIGESTIVO AlDIBL 96Aún no hay calificaciones
- Máquina de TintaDocumento1 páginaMáquina de TintaDIBL 96Aún no hay calificaciones
- JustificacionesDocumento1 páginaJustificacionesDIBL 96Aún no hay calificaciones
- PlasticocaptadorDocumento1 páginaPlasticocaptadorDIBL 96Aún no hay calificaciones
- Celula VegetalDocumento9 páginasCelula VegetaljoseAún no hay calificaciones
- Biología Introduccion A La CelulaDocumento7 páginasBiología Introduccion A La CelulaNataly CastroAún no hay calificaciones
- Biologia - Lleno (Pruebas Nacionales)Documento33 páginasBiologia - Lleno (Pruebas Nacionales)franklora599Aún no hay calificaciones
- MonografiaDocumento7 páginasMonografiaPABLO ULISES ROMERO MENDOZAAún no hay calificaciones
- GUÍA CelulaDocumento3 páginasGUÍA CelulaPatricio Bustamante SotoAún no hay calificaciones
- Temario: Exámenes Libres para Niños, Niñas Y Adolescentes 2023Documento20 páginasTemario: Exámenes Libres para Niños, Niñas Y Adolescentes 2023jessica lópezAún no hay calificaciones
- Semana 4 Naturales 4.2Documento5 páginasSemana 4 Naturales 4.2Solarte Toro MarianaAún no hay calificaciones
- Guía DE PRACTICA #2Documento12 páginasGuía DE PRACTICA #2miguel angelAún no hay calificaciones
- Diferencias Entre La Celula Animal y La Celula VegetalDocumento3 páginasDiferencias Entre La Celula Animal y La Celula VegetalPaul Andres Ruiz PantojaAún no hay calificaciones
- BIO2Documento4 páginasBIO2Leon LuisAún no hay calificaciones
- ArchivoDocumento13 páginasArchivoJose LFAún no hay calificaciones
- Balotario de Ciencia 1 UnidadDocumento6 páginasBalotario de Ciencia 1 UnidadJuan Avila AnastacioAún no hay calificaciones
- CELULADocumento12 páginasCELULAsilvia robayoAún no hay calificaciones
- Informe de Celulas VegetalesDocumento11 páginasInforme de Celulas VegetalesRaul Principe Ramos0% (1)
- Biolg - La CélulaDocumento40 páginasBiolg - La CélulaSergio Gonza LauraAún no hay calificaciones
- Taller Semana 4Documento7 páginasTaller Semana 4Luis Andres Parrales CarreñoAún no hay calificaciones
- Peroxisomas y VacuolasDocumento6 páginasPeroxisomas y VacuolasXxTAXIXLOLxXAún no hay calificaciones
- Reporte Practica de Laboratorio 1PDocumento18 páginasReporte Practica de Laboratorio 1PEmilianoCalixto Bonilla OsorioAún no hay calificaciones
- V ParenquimaDocumento15 páginasV ParenquimaAlanRocabadoAún no hay calificaciones
- Informe Practica 12Documento12 páginasInforme Practica 12Andres Gil100% (1)
- Evaluación Diagnóstica 2022 - 6to Grado-22-26Documento5 páginasEvaluación Diagnóstica 2022 - 6to Grado-22-26johanna damianAún no hay calificaciones
- Introduccion A La Biologia Celular CarpetaDocumento87 páginasIntroduccion A La Biologia Celular CarpetaLuisana y Mariela SalazarAún no hay calificaciones
- La CélulaDocumento3 páginasLa CélulaZamary OlayaAún no hay calificaciones
- Biología Celular y Molecular Practica (UPSJB-2016)Documento40 páginasBiología Celular y Molecular Practica (UPSJB-2016)Francesco Aguirre50% (2)
- La Celula y Su TamanoDocumento6 páginasLa Celula y Su TamanoElian LappostAún no hay calificaciones
- Planificacion Didactica NNKDocumento11 páginasPlanificacion Didactica NNKBrenda CastañoAún no hay calificaciones
- Evaluación Corte I Naturales 4°Documento4 páginasEvaluación Corte I Naturales 4°shazeniaAún no hay calificaciones
- Organelos Citoplasmaticos MaterialDocumento6 páginasOrganelos Citoplasmaticos Materialjohanna yanchatipanAún no hay calificaciones
- Teoria CelularDocumento10 páginasTeoria CelularJeraldin sanchezAún no hay calificaciones
- Actividad de Aprendizaje #18-Primer GradoDocumento4 páginasActividad de Aprendizaje #18-Primer GradoUltimate ArctiguanaAún no hay calificaciones