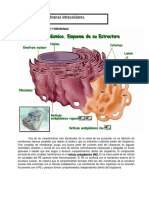
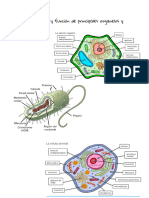
También podría gustarte
- Sistema Vacuolar CitoplasmáticoDocumento36 páginasSistema Vacuolar CitoplasmáticoAngie VillabonaAún no hay calificaciones
- Organizacion de Las CelulasDocumento9 páginasOrganizacion de Las CelulasEider Daniel Pastuzan DelgadoAún no hay calificaciones
- Organulos I. 1Documento2 páginasOrganulos I. 1victoriaAún no hay calificaciones
- Biologia Tema 6.1. EndomembranasDocumento24 páginasBiologia Tema 6.1. EndomembranasjorgeolivaressalidoAún no hay calificaciones
- Tema 8 Sistema de Endomembranas Reticulo EndoplasmaticoDocumento16 páginasTema 8 Sistema de Endomembranas Reticulo EndoplasmaticoValentina CelaniAún no hay calificaciones
- Célula Eucariota AnimalDocumento1 páginaCélula Eucariota AnimalKaren Jimenez100% (2)
- Bic U2 t3 PDF 18Documento5 páginasBic U2 t3 PDF 18bombitopazAún no hay calificaciones
- Procesos Biológicos de Jesús Mendoza Karen Célula Eucariota y Procariota Célula Eucariota Célula Procariota FunciónDocumento5 páginasProcesos Biológicos de Jesús Mendoza Karen Célula Eucariota y Procariota Célula Eucariota Célula Procariota FunciónKim JRAún no hay calificaciones
- Trabajo Biologia - Numero 2Documento15 páginasTrabajo Biologia - Numero 2Hernan Hernandez MontalvoAún no hay calificaciones
- Células Eucariotas y ProcariotasDocumento5 páginasCélulas Eucariotas y ProcariotasFanny CMunvAún no hay calificaciones
- Organulos Membranosos y No MembranososDocumento19 páginasOrganulos Membranosos y No MembranososJesus Antonio Sanchez TzintzunAún no hay calificaciones
- Organulos I. - 1Documento1 páginaOrganulos I. - 1Elias El AmraniAún no hay calificaciones
- Biologia CelularDocumento2 páginasBiologia CelularVeronica Guzman PeredoAún no hay calificaciones
- Celula AnimaDocumento5 páginasCelula Animapo poAún no hay calificaciones
- Separata Organelas y Componentes Extracelulares de Una Celula EucariotaDocumento9 páginasSeparata Organelas y Componentes Extracelulares de Una Celula Eucariotalucero liraAún no hay calificaciones
- Celula PrimeroDocumento15 páginasCelula PrimeroEricka Vizcarra BenavidesAún no hay calificaciones
- EucariotasDocumento49 páginasEucariotasMohamed AléAún no hay calificaciones
- Citoplasma y Organelas - Segundo Año.Documento6 páginasCitoplasma y Organelas - Segundo Año.estebanrodriguez9084uAún no hay calificaciones
- Unidad 5 Distribuciã - N de Proteã - NasDocumento10 páginasUnidad 5 Distribuciã - N de Proteã - NasAlexandra BlandónAún no hay calificaciones
- Diferencia Entre Célula Eucariota y ProcariotaDocumento11 páginasDiferencia Entre Célula Eucariota y ProcariotaLeydi Yudith Angarita Bautista89% (180)
- Tabla de Los Organelos C Animal y VegetalDocumento8 páginasTabla de Los Organelos C Animal y VegetalasliAún no hay calificaciones
- TE8 Los Organelos CitoplasmaticosDocumento21 páginasTE8 Los Organelos CitoplasmaticosFrancisco Reyes AnaluisaAún no hay calificaciones
- Célula Eucariota EVA1Documento4 páginasCélula Eucariota EVA1Cristobal Huerta100% (1)
- Organoides y Membranas IntracelularesDocumento10 páginasOrganoides y Membranas IntracelularesDIBL 96Aún no hay calificaciones
- Infografia BiologiaDocumento1 páginaInfografia BiologiaAlexandra RicseAún no hay calificaciones
- Citoplasma CelularDocumento19 páginasCitoplasma CelularAlisson Caicedo100% (2)
- 5) Celulas - Microbianas - EucariotasDocumento32 páginas5) Celulas - Microbianas - EucariotasJulian BarreraAún no hay calificaciones
- Unidad 2b Organelos y Trasporte EndomembranalDocumento19 páginasUnidad 2b Organelos y Trasporte EndomembranalArgeliaAún no hay calificaciones
- Unidad IDocumento31 páginasUnidad IErick CuauroAún no hay calificaciones
- Cuestionario Estructura Celular, Membranas y TransporteDocumento7 páginasCuestionario Estructura Celular, Membranas y TransporteloinconcienteAún no hay calificaciones
- Organelos Membranosoos y No MembranososDocumento7 páginasOrganelos Membranosoos y No MembranososMarjorie CampoverdeAún no hay calificaciones
- Organelos Celulares MembranososDocumento4 páginasOrganelos Celulares MembranososHANNIA PEREZ OVALLEAún no hay calificaciones
- Aporte de La Actividad Biologia MolecularDocumento10 páginasAporte de La Actividad Biologia MolecularFabian nunez cordobaAún no hay calificaciones
- 1Documento4 páginas1José Alejandro Valencia AlcaldeAún no hay calificaciones
- Tema 3 Estructura y Función CelularDocumento8 páginasTema 3 Estructura y Función CelularJorge GarcíaAún no hay calificaciones
- Las Células Proyecto Libro 2022Documento12 páginasLas Células Proyecto Libro 2022XWalterXAún no hay calificaciones
- Estructura y Función de Los Organelos CelularesDocumento10 páginasEstructura y Función de Los Organelos CelularesAnonymous AUC4kVqzAún no hay calificaciones
- Célula AnimalDocumento17 páginasCélula AnimalJUDITH ALONDRA TORRES HERNANDEZAún no hay calificaciones
- Wuolah Free App 1694792084005Documento95 páginasWuolah Free App 1694792084005Ángela PerezAún no hay calificaciones
- Album de BiologiaDocumento7 páginasAlbum de BiologiaNicole CallizoAún no hay calificaciones
- CélulaDocumento9 páginasCélulaAris CamarenaAún no hay calificaciones
- BiologíainformeDocumento30 páginasBiologíainformeclaudio ortgaAún no hay calificaciones
- Funcion de Estrcturas Celulares PDFDocumento2 páginasFuncion de Estrcturas Celulares PDFVictoria Nuñez VidalAún no hay calificaciones
- Estructura y Función CelularDocumento4 páginasEstructura y Función CelularCarolina OrtizAún no hay calificaciones
- MEMBRANAS INTERNAS 1 - TeoríaDocumento25 páginasMEMBRANAS INTERNAS 1 - TeoríaMarino GuerreroAún no hay calificaciones
- Biología: Tema 1: La CélulaDocumento10 páginasBiología: Tema 1: La Célulag3x ᴅᴍᴀᴋɪɪɪAún no hay calificaciones
- 03 Premedicina Biologia+Celular+y+Molecular Celula-Organelos+Celulares-transporte+MembranalDocumento9 páginas03 Premedicina Biologia+Celular+y+Molecular Celula-Organelos+Celulares-transporte+Membranalgeorgepiste1Aún no hay calificaciones
- U4 PP 84 Célula AnimalDocumento9 páginasU4 PP 84 Célula AnimalᗪᗴᑕKᗴᖇ乂Aún no hay calificaciones
- Resumes de Reticulos Endoplasmatico PDFDocumento1 páginaResumes de Reticulos Endoplasmatico PDFTirsa Xiomara Orozco CaalAún no hay calificaciones
- Cap. 12 Celula BiologiaDocumento54 páginasCap. 12 Celula BiologiaEmiliano FrancoAún no hay calificaciones
- Estructura Celular GeneralDocumento7 páginasEstructura Celular GeneralIvetteWininterOjedaAún no hay calificaciones
- Guia 1 Anatomia CelularDocumento10 páginasGuia 1 Anatomia CelularGloriaAún no hay calificaciones
- BIO - Sem4 - Citología IIDocumento10 páginasBIO - Sem4 - Citología IIgianella thirza salazar huamanAún no hay calificaciones
- La CélulaDocumento9 páginasLa CélulaPablo92% (37)
- Tema 6 Célula Eucariota Estructuras y FunciónDocumento22 páginasTema 6 Célula Eucariota Estructuras y FunciónFatima ArauzAún no hay calificaciones
- Cuadro de Organelos AnimalesDocumento1 páginaCuadro de Organelos AnimalesArkMarcerAún no hay calificaciones
- Histología y Embrología del Sitema NerviosoDe EverandHistología y Embrología del Sitema NerviosoCalificación: 4 de 5 estrellas4/5 (4)
- Acceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012De EverandAcceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012Aún no hay calificaciones
- Introducción a la Biología: RESÚMENES UNIVERSITARIOSDe EverandIntroducción a la Biología: RESÚMENES UNIVERSITARIOSCalificación: 5 de 5 estrellas5/5 (1)
- Celulas HumanasDocumento4 páginasCelulas HumanasDaniel ZambranoAún no hay calificaciones
- Tinciones FundamentosDocumento3 páginasTinciones Fundamentosprdzfonsi67% (3)
- Actividades CITOLOGIA AgronomiaDocumento4 páginasActividades CITOLOGIA AgronomiaFederico PorcelAún no hay calificaciones
- Mega Resumen Biología ComúnDocumento34 páginasMega Resumen Biología ComúnJosefina RojasAún no hay calificaciones
- 16 VirusDocumento7 páginas16 VirusJuana María Hernández HurtadoAún no hay calificaciones
- Algoritmo - Dogma CentralDocumento1 páginaAlgoritmo - Dogma CentralMateo VaronAún no hay calificaciones
- BiomoléculasDocumento43 páginasBiomoléculasjulia ubiergoAún no hay calificaciones
- UBA - CBC - Biologia - Resumen Libritos 2, 3, 4 y 6 - CatDocumento18 páginasUBA - CBC - Biologia - Resumen Libritos 2, 3, 4 y 6 - CatJuanJoseAún no hay calificaciones
- Diabetes MellitusDocumento12 páginasDiabetes MellitusSaul HernandezAún no hay calificaciones
- La CélulaDocumento5 páginasLa CélulaCarolAún no hay calificaciones
- Fisiopatología de La Demencia Degenerativas I - Dra. María Meza VegaDocumento32 páginasFisiopatología de La Demencia Degenerativas I - Dra. María Meza VegaANtonyMVAún no hay calificaciones
- HippoDocumento16 páginasHippoAndrea MorenoAún no hay calificaciones
- SEMINARIODocumento19 páginasSEMINARIOYorvin Juan Jose Montenegro RufastoAún no hay calificaciones
- ArnDocumento28 páginasArnSharini_Mu_oz_137Aún no hay calificaciones
- La SangreDocumento19 páginasLa SangreBruno LagioiaAún no hay calificaciones
- Vesículas TransportadorasDocumento7 páginasVesículas TransportadorasBeberly Reyes EspinozaAún no hay calificaciones
- Evaluación 2 Ciencias Naturales 8° Basico Membrana PlasmaticaDocumento8 páginasEvaluación 2 Ciencias Naturales 8° Basico Membrana Plasmaticahl l50% (2)
- Catabolismo LipidosDocumento60 páginasCatabolismo Lipidosyessenia apaza mamaniAún no hay calificaciones
- B54 - Ejercitación Guía 4Documento2 páginasB54 - Ejercitación Guía 4Martha CaceresAún no hay calificaciones
- La CelulaDocumento5 páginasLa CelulaNaju LauAún no hay calificaciones
- Diferencias Bacterias y ArchaeasDocumento7 páginasDiferencias Bacterias y ArchaeasAntonio Ulloa TorresAún no hay calificaciones
- Clases de Lipidos SaponificablesDocumento4 páginasClases de Lipidos SaponificablesDavid EnriquezAún no hay calificaciones
- 3ro Biología Diferenciado Guía 4 Unidad 0 Evaluación FormativaDocumento9 páginas3ro Biología Diferenciado Guía 4 Unidad 0 Evaluación FormativaBruno AllianaAún no hay calificaciones
- Las Implicaciones de La Hipermutacion SomaticaDocumento6 páginasLas Implicaciones de La Hipermutacion SomaticaGianella PachecoAún no hay calificaciones
- Regulación de La Expresión GénicaDocumento37 páginasRegulación de La Expresión GénicaMario Aarón Vásquez FabiánAún no hay calificaciones
- Evaluación de Eproteinas, Carbohidratos y LípidosDocumento2 páginasEvaluación de Eproteinas, Carbohidratos y LípidosMaria Solano0% (1)
- Diagnóstico Histológico 1Documento153 páginasDiagnóstico Histológico 1Camila Rivas DíazAún no hay calificaciones
- Guion p6Documento5 páginasGuion p6Laura Gomez AnzaldoAún no hay calificaciones
- HeridasDocumento88 páginasHeridasdenisse AlcazarAún no hay calificaciones
- Resultados EPS PDFDocumento3 páginasResultados EPS PDFdiana urregoAún no hay calificaciones