También podría gustarte
- Aproximación A La Composición de La Macrofauna Edáfica Asociada A Una Franja de Bosque AndinoDocumento6 páginasAproximación A La Composición de La Macrofauna Edáfica Asociada A Una Franja de Bosque AndinoPedro RuedaAún no hay calificaciones
- Preferencia Alimentaria de Neochetina Eichhorniae Coleoptera CurculionidaeDocumento7 páginasPreferencia Alimentaria de Neochetina Eichhorniae Coleoptera Curculionidaemeli15Aún no hay calificaciones
- Muestreo de Algunas Especies de Serpientes en La Isla de Gorgona y GorgonillaDocumento16 páginasMuestreo de Algunas Especies de Serpientes en La Isla de Gorgona y GorgonillaJuan Camilo BojacàAún no hay calificaciones
- EVALUACIÓN DE FLUJO Y VARIABILIDAD GENÉTICA DE LA RANA RUBÍ (Dendrobates Bombetes) EN LAS RESERVAS DE YOTOCO (VALLE DEL CAUCA, COLOMBIA) Y BREMEN (QUINDIO, COLOMBIA)Documento22 páginasEVALUACIÓN DE FLUJO Y VARIABILIDAD GENÉTICA DE LA RANA RUBÍ (Dendrobates Bombetes) EN LAS RESERVAS DE YOTOCO (VALLE DEL CAUCA, COLOMBIA) Y BREMEN (QUINDIO, COLOMBIA)Juan Camilo Martinez BarguilAún no hay calificaciones
- CONICET Digital Nro - Db87a51b 8510 4ffd Bf85 96db2406cea4 ADocumento17 páginasCONICET Digital Nro - Db87a51b 8510 4ffd Bf85 96db2406cea4 AMila ChapasAún no hay calificaciones
- CONICET Digital Nrofd62eb00-Acd8-4fa7-9682-98aca7 221006 194128Documento35 páginasCONICET Digital Nrofd62eb00-Acd8-4fa7-9682-98aca7 221006 194128marco arroyo sotoAún no hay calificaciones
- Proyecto Microhabitat de R. Sirensis en BambuDocumento33 páginasProyecto Microhabitat de R. Sirensis en BambuFiorella Medina MalquiAún no hay calificaciones
- Perú, megadiverso país con gran riqueza de especiesDocumento1 páginaPerú, megadiverso país con gran riqueza de especiesstefano huacarpuma ayuquiAún no hay calificaciones
- Pleurotus Djamor, Un Hongo Con Potencial Aplicación Biotecnológica para El NeotrópicoDocumento13 páginasPleurotus Djamor, Un Hongo Con Potencial Aplicación Biotecnológica para El NeotrópicoPauly Campos OyarzúnAún no hay calificaciones
- Día 2. Escarabajos (Coleoptera - Scarabaeidae) Como Bioindicadores y Remediadores de Ecosistemas Naturales y AgrícolasDocumento46 páginasDía 2. Escarabajos (Coleoptera - Scarabaeidae) Como Bioindicadores y Remediadores de Ecosistemas Naturales y AgrícolasPablo Romero MejiaAún no hay calificaciones
- Nymphalidae FrugivorasDocumento14 páginasNymphalidae Frugivoraslv acAún no hay calificaciones
- Diversidad y funcionalidad de la macrofauna edáficaDocumento31 páginasDiversidad y funcionalidad de la macrofauna edáficaMiguel AngelAún no hay calificaciones
- 880-Texto Del Manuscrito-2653-2-10-20210310Documento12 páginas880-Texto Del Manuscrito-2653-2-10-20210310series novelasAún no hay calificaciones
- BorradorDocumento3 páginasBorradorBrigith quirogaAún no hay calificaciones
- Nematodos en El ArrozDocumento5 páginasNematodos en El ArrozmanoloAún no hay calificaciones
- La Importancia de Los Manglares y Su Microbiología (Cangrejos)Documento17 páginasLa Importancia de Los Manglares y Su Microbiología (Cangrejos)elliottAún no hay calificaciones
- Flora y Fauna Del Bosque Seco de La Provincia de Loja EcuadorDocumento104 páginasFlora y Fauna Del Bosque Seco de La Provincia de Loja EcuadorJuliaLojánAún no hay calificaciones
- Libélulas y caballitos del diablo del departamento del Meta, ColombiaDe EverandLibélulas y caballitos del diablo del departamento del Meta, ColombiaAún no hay calificaciones
- CARBONO ContentServerDocumento11 páginasCARBONO ContentServerjhonerAún no hay calificaciones
- Flora de Los Afloramientos de Marmoles yDocumento35 páginasFlora de Los Afloramientos de Marmoles yGabrielPedrazziAún no hay calificaciones
- 64-Texto Del Artículo-216-1-10-20200225Documento12 páginas64-Texto Del Artículo-216-1-10-20200225kelly baqueroAún no hay calificaciones
- Tesis Moluscos Gasterópodos Bahia de Ancon PDFDocumento46 páginasTesis Moluscos Gasterópodos Bahia de Ancon PDFJavier Fernando Rios OrihuelaAún no hay calificaciones
- Fase 1 - 302277025 - 35Documento8 páginasFase 1 - 302277025 - 35zharick GarcíaAún no hay calificaciones
- Mancina Diversidadcuba FaunadelSuelo PDFDocumento34 páginasMancina Diversidadcuba FaunadelSuelo PDFDaline RiosAún no hay calificaciones
- 2016-Murillo-Ramos Et Al.-Ecologia AustralDocumento10 páginas2016-Murillo-Ramos Et Al.-Ecologia AustralGylbert RiveraAún no hay calificaciones
- 6 Diversidad y Rol Funcional de La Macrofauna EdaficaDocumento31 páginas6 Diversidad y Rol Funcional de La Macrofauna Edaficajulio.nunez2901Aún no hay calificaciones
- Diversidad y Distribucion Vertical de EpDocumento134 páginasDiversidad y Distribucion Vertical de Epmcsc337799Aún no hay calificaciones
- Composition of The Biofouling Community Associated With Oyster Culture in An Amazon Estuary, Pará State, North BrazilDocumento9 páginasComposition of The Biofouling Community Associated With Oyster Culture in An Amazon Estuary, Pará State, North BrazilCecili MendesAún no hay calificaciones
- Revista Comunicaciones Cientificas y Tecnologicas Vol 5 No 1 Diciembre 2019 Art5Documento21 páginasRevista Comunicaciones Cientificas y Tecnologicas Vol 5 No 1 Diciembre 2019 Art5Kerlin Marbella Saavedra ShanocuaAún no hay calificaciones
- Sobreposición de dieta entre aves y murciélagos frugívoros en islas de bosqueDocumento15 páginasSobreposición de dieta entre aves y murciélagos frugívoros en islas de bosqueHenry CondoriAún no hay calificaciones
- Ensamble Ficoperifitico Asociado A Macrofitas en UDocumento9 páginasEnsamble Ficoperifitico Asociado A Macrofitas en Uavaron21Aún no hay calificaciones
- Cariassanchez,+339650 Texto+del+artículo 199647 7 10 20210129 - CompressedDocumento13 páginasCariassanchez,+339650 Texto+del+artículo 199647 7 10 20210129 - Compressedandresg.ruedaAún no hay calificaciones
- Zuleyma Mosquera-Murillo, María Ángela Gómez, Maira Beatriz GonzálezDocumento7 páginasZuleyma Mosquera-Murillo, María Ángela Gómez, Maira Beatriz GonzálezVERENICE CEBOLLERO-JUARBEAún no hay calificaciones
- Colepteros NecrócolosDocumento7 páginasColepteros NecrócolosJuan Jose SotoAún no hay calificaciones
- Guía - Salida Tecnica en Campo, CEA.Documento3 páginasGuía - Salida Tecnica en Campo, CEA.johnAún no hay calificaciones
- Biodiversidad6 UsosDocumento103 páginasBiodiversidad6 UsosMendiola DanAún no hay calificaciones
- Bio Divers I DadDocumento16 páginasBio Divers I DadIsamar ORAún no hay calificaciones
- Composicion Espacial y Temporal de Aves AcuaticasDocumento18 páginasComposicion Espacial y Temporal de Aves AcuaticasmariaAún no hay calificaciones
- Dieta Mur VenezuelaDocumento11 páginasDieta Mur VenezuelaFabrizio VarelaAún no hay calificaciones
- Dalbergia CongestifloraDocumento5 páginasDalbergia CongestifloraDeeysii de la CruzAún no hay calificaciones
- Rabia transmitida por murciélagos BrasilDocumento16 páginasRabia transmitida por murciélagos BrasilÁlvaro CarrasquelAún no hay calificaciones
- Uso de Especies de Plantas en El Chocó BiogeográficoDocumento27 páginasUso de Especies de Plantas en El Chocó BiogeográficokritzziaAún no hay calificaciones
- Revista de Biología Tropical 0034-7744: Issn: Rbt@cariari - Ucr.ac - CRDocumento11 páginasRevista de Biología Tropical 0034-7744: Issn: Rbt@cariari - Ucr.ac - CROsmar IralaAún no hay calificaciones
- Biodiversidad Del Ecuador TripticoDocumento4 páginasBiodiversidad Del Ecuador TripticoEnriqueAún no hay calificaciones
- Lectura de ArtropodosDocumento7 páginasLectura de ArtropodosAna EscobarAún no hay calificaciones
- Practica 03 BiodiversidadDocumento6 páginasPractica 03 BiodiversidadLUIS FERNANDO VASQUEZ SANCHEZAún no hay calificaciones
- Diversidad quiróptera cultivos EcuadorDocumento1 páginaDiversidad quiróptera cultivos EcuadorLEMA HERNANDEZ ALICIA FERNANDAAún no hay calificaciones
- C. Mulsant - SRJDocumento3 páginasC. Mulsant - SRJSalomón RamírezAún no hay calificaciones
- Mariposas Atlantico Boletin CaldasDocumento17 páginasMariposas Atlantico Boletin CaldasFabricio Murrieta ChotaAún no hay calificaciones
- Lepidopteras PDFDocumento34 páginasLepidopteras PDFRodrigo Adrian RubioAún no hay calificaciones
- Biodiversidad, Clasificación y FilogeniaDocumento17 páginasBiodiversidad, Clasificación y FilogeniaYessenia OropezaAún no hay calificaciones
- Biología de Las Moscas CalliphoridaeDocumento8 páginasBiología de Las Moscas CalliphoridaeRafael ÁvilaAún no hay calificaciones
- Diversidad de Invertebrados Consumidos Por Las Etnias Yanomami y Yekuana Del Alto Orinoco, VEN PDFDocumento6 páginasDiversidad de Invertebrados Consumidos Por Las Etnias Yanomami y Yekuana Del Alto Orinoco, VEN PDFDaniela Rojas MantillaAún no hay calificaciones
- 9 Diversidad y Manejo de Los Termes de Mexico PDFDocumento13 páginas9 Diversidad y Manejo de Los Termes de Mexico PDFHiginio Galindo Estudillo GaeshAún no hay calificaciones
- Insect Oster Rest ResDocumento32 páginasInsect Oster Rest ResCandela GaunaAún no hay calificaciones
- Percepciones Murciélagos Atahualpa (Habaspamba) - 2022Documento15 páginasPercepciones Murciélagos Atahualpa (Habaspamba) - 2022Anthony AguilarAún no hay calificaciones
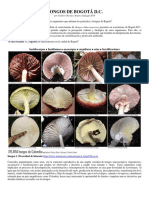
- Hongos en Humedales Cerros y Páramos de Bogotá, 2018Documento6 páginasHongos en Humedales Cerros y Páramos de Bogotá, 2018Teodoro Chivatá BedoyaAún no hay calificaciones
- Articulo Final V2 EcologiaDocumento12 páginasArticulo Final V2 Ecologiajacquelinetrejoc0110Aún no hay calificaciones
- Yoshimoto Etal 2021 RMBDocumento13 páginasYoshimoto Etal 2021 RMBmaewagyuchoAún no hay calificaciones
- Catálogo de los copépodos (Crustacea: Copepoda: Calanoida y Cyclopoida) de cuerpos de agua temporales de Jalisco, México: Taxonomía y distribuciónDe EverandCatálogo de los copépodos (Crustacea: Copepoda: Calanoida y Cyclopoida) de cuerpos de agua temporales de Jalisco, México: Taxonomía y distribuciónAún no hay calificaciones
- Z4 Luis Sorio y Compañia Efecto Fundador en Un Canido ESDocumento30 páginasZ4 Luis Sorio y Compañia Efecto Fundador en Un Canido ESLuis OsorioAún no hay calificaciones
- Farji BrenerAG 2022 EcoAustral Hipotesis EnprensaDocumento11 páginasFarji BrenerAG 2022 EcoAustral Hipotesis EnprensaLuis OsorioAún no hay calificaciones
- Uso Correcto, Parcial e Incorrecto de Los Términos "Hipótesis" y "Predicciones" en EcologíaDocumento6 páginasUso Correcto, Parcial e Incorrecto de Los Términos "Hipótesis" y "Predicciones" en EcologíaLuis OsorioAún no hay calificaciones
- GUÍA No 4. CAPACITOR DE PLACAS PLANAS Y PARALELAS 2Documento7 páginasGUÍA No 4. CAPACITOR DE PLACAS PLANAS Y PARALELAS 2Luis OsorioAún no hay calificaciones
- Evolución de La Morfología Genital en InsectosDocumento17 páginasEvolución de La Morfología Genital en InsectosLuis OsorioAún no hay calificaciones
- Capacitor Placas Planas y Paralelas Final Hasta MetDocumento6 páginasCapacitor Placas Planas y Paralelas Final Hasta MetLuis OsorioAún no hay calificaciones
- Guía Laboratorio ElectroestáticaDocumento10 páginasGuía Laboratorio ElectroestáticaLuis OsorioAún no hay calificaciones
- Article: ZootaxaDocumento14 páginasArticle: ZootaxaLuis OsorioAún no hay calificaciones
- Guía No 0. Introducción A Las Mediciones EléctricasDocumento5 páginasGuía No 0. Introducción A Las Mediciones EléctricasKeiidys MartinezAún no hay calificaciones
- Estrada Alvarez Guadarrama & Martinez 2013 Dug 20 63 66Documento4 páginasEstrada Alvarez Guadarrama & Martinez 2013 Dug 20 63 66Luis OsorioAún no hay calificaciones
- Circuitos Eléctricos Visión GeneralDocumento6 páginasCircuitos Eléctricos Visión GeneralAlfons7Aún no hay calificaciones
- Universidad Del Atlantico Taller Distribuciones ContinuasDocumento3 páginasUniversidad Del Atlantico Taller Distribuciones ContinuasLaura PosadaAún no hay calificaciones
- Capacitor de placas planasDocumento5 páginasCapacitor de placas planasLuis OsorioAún no hay calificaciones
- Guía introductoria a mediciones eléctricasDocumento6 páginasGuía introductoria a mediciones eléctricasLuis OsorioAún no hay calificaciones
- Campos magnéticos, fuerzas entre corrientes y fem inducidaDocumento3 páginasCampos magnéticos, fuerzas entre corrientes y fem inducidaLuis OsorioAún no hay calificaciones
- Diseño Multimetro AnalogicoDocumento26 páginasDiseño Multimetro AnalogicoR.l. Rico100% (2)
- Circuitos eléctricos-Taller prácticoDocumento3 páginasCircuitos eléctricos-Taller prácticoLuis OsorioAún no hay calificaciones
- Taller 2Documento1 páginaTaller 2Luis OsorioAún no hay calificaciones
- Taller - Fuerza MagneticaDocumento2 páginasTaller - Fuerza MagneticaLuis OsorioAún no hay calificaciones
- Taller - FUERZAS SOBRE CORRIENTESDocumento1 páginaTaller - FUERZAS SOBRE CORRIENTESLuis OsorioAún no hay calificaciones
- Campos magnéticos, fuerzas entre corrientes y fem inducidaDocumento3 páginasCampos magnéticos, fuerzas entre corrientes y fem inducidaLuis OsorioAún no hay calificaciones
- EQUINODERMOSDocumento3 páginasEQUINODERMOSLuis OsorioAún no hay calificaciones
- Código de DatosDocumento2 páginasCódigo de DatosLuis OsorioAún no hay calificaciones
- EQUINODERMOSDocumento3 páginasEQUINODERMOSLuis OsorioAún no hay calificaciones
- BryozoaDocumento1 páginaBryozoaLuis OsorioAún no hay calificaciones
- 1° Tarea Teoría GesellDocumento5 páginas1° Tarea Teoría GesellRoger Mendoza LimaAún no hay calificaciones
- Reglamentación de las organizaciones solidarias en ColombiaDocumento8 páginasReglamentación de las organizaciones solidarias en Colombiamanuel carrilloAún no hay calificaciones
- ¿Qué Significa PensarDocumento2 páginas¿Qué Significa PensarAgustina PazAún no hay calificaciones
- ProcesosDocumento4 páginasProcesosjavier rodriguezAún no hay calificaciones
- 2 Com. Narram. Historia - SesiónDocumento2 páginas2 Com. Narram. Historia - SesiónAlexandra Iglecias Rioja100% (1)
- Región Andina de Colombia, geografía y economíaDocumento26 páginasRegión Andina de Colombia, geografía y economíaJesus Pardo YanezAún no hay calificaciones
- Orientadas A Las Mensuras - GeodesiaDocumento21 páginasOrientadas A Las Mensuras - GeodesiaHugo VentimigliaAún no hay calificaciones
- Syllabus Calculo Vectorial 3cre 2021-1Documento4 páginasSyllabus Calculo Vectorial 3cre 2021-1laura tatiana espitia pinillaAún no hay calificaciones
- Secdoc04 Libros Eso DefinDocumento3 páginasSecdoc04 Libros Eso DefinFco Javier Navas GómezAún no hay calificaciones
- Análisis de refrigerantes para mejorar transferencia de calorDocumento2 páginasAnálisis de refrigerantes para mejorar transferencia de calorJuan Sebastian Ramirez MartinezAún no hay calificaciones
- Actividad 13 UD 2 Samuel PuertaDocumento8 páginasActividad 13 UD 2 Samuel PuertaSamuel Puerta MerinoAún no hay calificaciones
- P1 Mate 4 Actividad 4Documento3 páginasP1 Mate 4 Actividad 4AskarAún no hay calificaciones
- 18 Operaciones IntelectualesDocumento8 páginas18 Operaciones IntelectualesAnder NarvAún no hay calificaciones
- Estrategias de negocio, fortalezas y debilidadesDocumento6 páginasEstrategias de negocio, fortalezas y debilidadesyefrin salazarAún no hay calificaciones
- Linea de TiempoDocumento2 páginasLinea de TiempoSamuel MendozaAún no hay calificaciones
- Actividad No - 3 (Transferencia)Documento14 páginasActividad No - 3 (Transferencia)GuillermoAún no hay calificaciones
- Lab 1 Teoria ErroresDocumento36 páginasLab 1 Teoria ErroresFamilia Ríos PeredaAún no hay calificaciones
- 4-Sarni y Corbo - La Evaluacio769n en Educacio769n Fi769sica Ma769s Alla769 Del Control SocialDocumento17 páginas4-Sarni y Corbo - La Evaluacio769n en Educacio769n Fi769sica Ma769s Alla769 Del Control SocialAgustin AraAún no hay calificaciones
- 1.1 Datos Adicionales RequeridosDocumento5 páginas1.1 Datos Adicionales RequeridosMariuxi Rocío Fajardo GuamanAún no hay calificaciones
- Benidorm Calendario Escolar Curs 2018 - 2019Documento1 páginaBenidorm Calendario Escolar Curs 2018 - 2019RACHELAún no hay calificaciones
- SENTENCIASDocumento5 páginasSENTENCIASMarcos GaiborAún no hay calificaciones
- MacroentornoDocumento10 páginasMacroentornoanon_473926246100% (1)
- Centroides y Centros de Gravedad Placas-Compuestas 24Documento6 páginasCentroides y Centros de Gravedad Placas-Compuestas 24Estefany AzulaAún no hay calificaciones
- Mapas TopográficosDocumento4 páginasMapas TopográficosyosswanAún no hay calificaciones
- TP Escuela NuevaDocumento7 páginasTP Escuela NuevaRooAún no hay calificaciones
- Karen - Palacios - Paredes - Control - s3Documento8 páginasKaren - Palacios - Paredes - Control - s3Karen PalaciosAún no hay calificaciones
- Informe Técnico Asesoría en Actualización de Matriz de PeligrosDocumento12 páginasInforme Técnico Asesoría en Actualización de Matriz de PeligrosANGELA SANTANAAún no hay calificaciones
- Informe Encuesta COVID-19Documento7 páginasInforme Encuesta COVID-19Urgente24Aún no hay calificaciones
- Matriz C.I Colombia ParadiseDocumento249 páginasMatriz C.I Colombia ParadiseCINDY MARCELA SALGUERO DONCELAún no hay calificaciones
- PRESENTACIÒNDocumento34 páginasPRESENTACIÒNGuissela Rosibel Moreno LeónAún no hay calificaciones