También podría gustarte
- Sem7 ApoptosisDocumento14 páginasSem7 ApoptosisArnaldo PérezAún no hay calificaciones
- Necrosis neuronal programadaDocumento6 páginasNecrosis neuronal programadaBryam LuceroAún no hay calificaciones
- La Apoptosis en La Biologia y PatologiaDocumento10 páginasLa Apoptosis en La Biologia y PatologiaVictoria VillaAún no hay calificaciones
- Apoptosis: muerte celular programadaDocumento7 páginasApoptosis: muerte celular programadaClaw JuárezAún no hay calificaciones
- Proyecto ApoptosisDocumento32 páginasProyecto ApoptosisJesúsCoylaAún no hay calificaciones
- Tipos de Muerte Celular y Sus Implicaciones ClínicasDocumento18 páginasTipos de Muerte Celular y Sus Implicaciones ClínicasCosa SuarezAún no hay calificaciones
- Bloque III Tema2Documento14 páginasBloque III Tema2Carlos Orozco GonzalezAún no hay calificaciones
- Historia de la muerte celularDocumento16 páginasHistoria de la muerte celularangie catalina estupiñan mirandaAún no hay calificaciones
- ApóptosisDocumento6 páginasApóptosisMarisol MoralesAún no hay calificaciones
- La Apoptosis en El CáncerDocumento10 páginasLa Apoptosis en El CáncerUSMP FN ARCHIVOS100% (1)
- Lesion Celular y NecrosisDocumento3 páginasLesion Celular y NecrosisJessica Duque CorreaAún no hay calificaciones
- Necroptosis en DesarrolloDocumento11 páginasNecroptosis en Desarrolloluis mendozaAún no hay calificaciones
- ApoptosisDocumento27 páginasApoptosisJose LuisAún no hay calificaciones
- Apoptosis PDFDocumento7 páginasApoptosis PDFYanita Avila ClazngAún no hay calificaciones
- Apoptosis Muerte Celular FisiológicaDocumento8 páginasApoptosis Muerte Celular FisiológicaCAMILA ROZOAún no hay calificaciones
- SEMINARIODocumento4 páginasSEMINARIOMilagros Blas PizarroAún no hay calificaciones
- Apoptosis Del EritrocitoDocumento5 páginasApoptosis Del EritrocitoanaranabyAún no hay calificaciones
- Informe 5 Ep023-2 Biologia SemDocumento8 páginasInforme 5 Ep023-2 Biologia SemGrethell MedinaAún no hay calificaciones
- GUION BiologiaDocumento5 páginasGUION BiologiaNahomy BustillosAún no hay calificaciones
- Diapositivas de Apoptosis FinalDocumento59 páginasDiapositivas de Apoptosis FinalLeslie Zavaleta Bazan100% (1)
- ApoptosisDocumento14 páginasApoptosisDarleny AcostaAún no hay calificaciones
- Muerte CelularDocumento5 páginasMuerte CelularGael GilAún no hay calificaciones
- Tarea 7 APOPTOSIS - Docx TRABAJO GRUPAL CORREGIDO (Vanessa Lucia Gutierrez Bossio)Documento12 páginasTarea 7 APOPTOSIS - Docx TRABAJO GRUPAL CORREGIDO (Vanessa Lucia Gutierrez Bossio)Vanessa GutierrezAún no hay calificaciones
- Tarea 4 Elida Casas 155t4Documento12 páginasTarea 4 Elida Casas 155t4Armando Olave ValenciaAún no hay calificaciones
- Apoptosis: NombresDocumento20 páginasApoptosis: Nombresjaime alfonso conde monteAún no hay calificaciones
- Semana 9 - Seminario - Lectura - La Vida Tras La MuerteDocumento9 páginasSemana 9 - Seminario - Lectura - La Vida Tras La MuertejuanAún no hay calificaciones
- Muerte Celular Programada Bacteriana - IluminadoDocumento17 páginasMuerte Celular Programada Bacteriana - IluminadoDaniel HenaoAún no hay calificaciones
- Preguntas Finales ArticuloDocumento8 páginasPreguntas Finales ArticulojosearmandoAún no hay calificaciones
- UntitledDocumento11 páginasUntitledpgisbelAún no hay calificaciones
- Portafolio de LaboratorioDocumento96 páginasPortafolio de LaboratorioSoli Deo GloriaAún no hay calificaciones
- APOPTOSISDocumento21 páginasAPOPTOSISjaime alfonso conde monteAún no hay calificaciones
- Apoptosis: Implicaciones en Medicina Intensiva: RevisiónDocumento10 páginasApoptosis: Implicaciones en Medicina Intensiva: RevisiónNIDIA CATERIN MARTINEZ BONILLAAún no hay calificaciones
- La muerte celular programada: apoptosisDocumento24 páginasLa muerte celular programada: apoptosiskarlosAún no hay calificaciones
- Apoptosis TrabajoDocumento30 páginasApoptosis TrabajoJennifer SamanthaAún no hay calificaciones
- Apoptosis y NecrosisDocumento1 páginaApoptosis y NecrosisVALERIA SANCHEZAún no hay calificaciones
- Articulo BiologiaDocumento6 páginasArticulo BiologiaJodie LeónAún no hay calificaciones
- Apoptosis PDFDocumento6 páginasApoptosis PDFFco Javier Sanchez PeñaAún no hay calificaciones
- Muerte Celular y NecrosisDocumento114 páginasMuerte Celular y NecrosisAndië ÄndreäAún no hay calificaciones
- Apoptosis y Muerte Celular-1Documento12 páginasApoptosis y Muerte Celular-1catalinaAún no hay calificaciones
- Análisis de la muerte celular programada y la genética de MendelDocumento13 páginasAnálisis de la muerte celular programada y la genética de MendelleidyAún no hay calificaciones
- ApoptosisDocumento14 páginasApoptosismichisstrokesAún no hay calificaciones
- Mecanismos Moleculares de Muerte Celular.Documento39 páginasMecanismos Moleculares de Muerte Celular.Xaira RicoAún no hay calificaciones
- Informe Biologia Sem s5Documento5 páginasInforme Biologia Sem s5alexandra carpioAún no hay calificaciones
- Apotosis y NecrosisDocumento11 páginasApotosis y NecrosisEvert BonifacioAún no hay calificaciones
- Apoptosis CelularDocumento12 páginasApoptosis CelularJean Paul GonzalesAún no hay calificaciones
- DefinicionesDocumento5 páginasDefinicionesNora VelandiaAún no hay calificaciones
- ApoptosisDocumento5 páginasApoptosisLuis Noriega RiveraAún no hay calificaciones
- ArticuloDocumento9 páginasArticuloFuentes Perez ItzelAún no hay calificaciones
- Mecanismos de Muerte CelularDocumento11 páginasMecanismos de Muerte CelularJANELA KATHERINE MUÑOZ ROSASAún no hay calificaciones
- Muerte Celular PDocumento47 páginasMuerte Celular Pnatalia.perez.camigliaAún no hay calificaciones
- Muerte Celular Ensayo ArgumentativoDocumento7 páginasMuerte Celular Ensayo ArgumentativoROXY FERNANDA RODRÍGUEZ TORRESAún no hay calificaciones
- Bases Moleculares de La Apoptosis: Anal. Real Acad. Nal. Farm., 2003, 69Documento29 páginasBases Moleculares de La Apoptosis: Anal. Real Acad. Nal. Farm., 2003, 69María isabel Guzmán fernandezAún no hay calificaciones
- Formas de Muerte CelularDocumento4 páginasFormas de Muerte CelularLAURA VICTORIA OAXACA NEVAREZAún no hay calificaciones
- ApoptosisDocumento10 páginasApoptosisPeri-Hala Norbibi Monago SantosAún no hay calificaciones
- Fundamentos y técnicas básicas en biología molecularDe EverandFundamentos y técnicas básicas en biología molecularAún no hay calificaciones
- Psicooncología: Ayuda Psicológica Para Pacientes Con CáncerDe EverandPsicooncología: Ayuda Psicológica Para Pacientes Con CáncerCalificación: 4 de 5 estrellas4/5 (1)
- Filosofía del cáncer: Una perspectiva dinámica y relacionalDe EverandFilosofía del cáncer: Una perspectiva dinámica y relacionalCalificación: 5 de 5 estrellas5/5 (1)
- Guía Paratiroides y Homeostasis Del CalcioDocumento5 páginasGuía Paratiroides y Homeostasis Del CalcioKristy RodriguezAún no hay calificaciones
- Documento DeEnfermedad Renal Diabética - María Andrea RuizDocumento25 páginasDocumento DeEnfermedad Renal Diabética - María Andrea RuizMaria RuizAún no hay calificaciones
- Bioazufre FichaDocumento1 páginaBioazufre FichaJorge TorósAún no hay calificaciones
- Parto, Puerperio y LactanciaDocumento39 páginasParto, Puerperio y LactanciaYeramiel PérezAún no hay calificaciones
- Tratamiento de arritmias gravesDocumento49 páginasTratamiento de arritmias gravesNadia RojasAún no hay calificaciones
- Caso ClínicoDocumento18 páginasCaso ClínicoRicardo100% (1)
- Tamizaje Neonatal 2023Documento37 páginasTamizaje Neonatal 2023César RiveraAún no hay calificaciones
- Monografias AnestesicosDocumento30 páginasMonografias Anestesicospepe lepu100% (1)
- Dietas Bajas en Carbohidratos y Cetogénicas en La Diabetes Tipo 1 y Tipo 2Documento14 páginasDietas Bajas en Carbohidratos y Cetogénicas en La Diabetes Tipo 1 y Tipo 2Carol EstefaníaAún no hay calificaciones
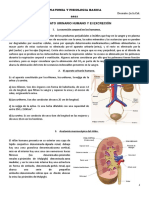
- MOD VI - El APARATO URINARIO HUMANO Y El EXCRECIÓNDocumento6 páginasMOD VI - El APARATO URINARIO HUMANO Y El EXCRECIÓNjaviermagicAún no hay calificaciones
- Resumen Nutrición Animal FAUBADocumento16 páginasResumen Nutrición Animal FAUBAAgus NocerezAún no hay calificaciones
- Métodos Anticonceptivos Hormonales - 20230926 - 080830 - 0000Documento3 páginasMétodos Anticonceptivos Hormonales - 20230926 - 080830 - 0000luchiiarguello05Aún no hay calificaciones
- Perfil LipidicoDocumento1 páginaPerfil LipidicoSam WillyAún no hay calificaciones
- 7 GuiaDocumento112 páginas7 GuiaAna BelenAún no hay calificaciones
- DIABETESDocumento28 páginasDIABETESNatalia De PaulaAún no hay calificaciones
- Exámenes Auxiliares - Aga PDFDocumento22 páginasExámenes Auxiliares - Aga PDFAngie FiorellaAún no hay calificaciones
- Resultados CHECK-UPDocumento6 páginasResultados CHECK-UPSeiji Nakashima100% (2)
- Quiz Equilibrio Acido BasicoDocumento6 páginasQuiz Equilibrio Acido BasicoScarleth MunguiaAún no hay calificaciones
- Cuidados Del Sistema MuscularDocumento3 páginasCuidados Del Sistema MuscularCL CarmenAún no hay calificaciones
- Miocardiopatia RestrictivaDocumento13 páginasMiocardiopatia RestrictivaPilar GuillénAún no hay calificaciones
- 5 - Vitaminas Esposicion, EvelynDocumento24 páginas5 - Vitaminas Esposicion, Evelynmaribel rodriguezAún no hay calificaciones
- 7-Salud Guía 3Documento6 páginas7-Salud Guía 3Yeidys Vanessa Peña CastroAún no hay calificaciones
- 9 Metabolismo de Las ProteínasDocumento10 páginas9 Metabolismo de Las ProteínasMervin Arauz HoyosAún no hay calificaciones
- Exámenes de SangreDocumento4 páginasExámenes de SangreRune OstherusAún no hay calificaciones
- Guía de Patología General 2020-Ii 2Documento61 páginasGuía de Patología General 2020-Ii 2dennisAún no hay calificaciones
- Examen electrolitos líquidosDocumento3 páginasExamen electrolitos líquidosEmanuel Castro100% (6)
- Cetoacidosis Diabética 1Documento2 páginasCetoacidosis Diabética 1AlegnaAún no hay calificaciones
- Minerales Quimico de Alimentos TRABAJO FINALDocumento16 páginasMinerales Quimico de Alimentos TRABAJO FINALYunihor PM JunihorAún no hay calificaciones
- 25 HormonasDocumento9 páginas25 Hormonaseric ortigozaAún no hay calificaciones