También podría gustarte
- Ecografía normal del árbol urinario y genitales externosDe EverandEcografía normal del árbol urinario y genitales externosAún no hay calificaciones
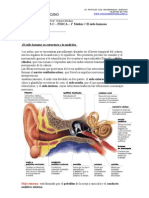
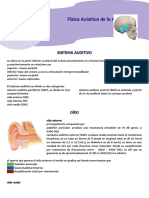
- Resumen Sentido de La Audicion Cap. 53Documento5 páginasResumen Sentido de La Audicion Cap. 53nena mish100% (1)
- OidoDocumento19 páginasOidoDavid GutiérrezAún no hay calificaciones
- OtorrinoDocumento52 páginasOtorrinoEussk AdyAún no hay calificaciones
- Macro Destrezas Del LenguajeDocumento9 páginasMacro Destrezas Del LenguajeOswaldoMiño0% (1)
- Trauma AcusticoDocumento46 páginasTrauma AcusticoMiguel Angel RazoAún no hay calificaciones
- MicrotiaDocumento11 páginasMicrotiaJoselina Pilataxi ValenciaAún no hay calificaciones
- Resonancia Magnetica de OidoDocumento22 páginasResonancia Magnetica de OidoMARY LUZ SANCHEZAún no hay calificaciones
- 4 Fisiología Auditiva PDFDocumento62 páginas4 Fisiología Auditiva PDFflga. katherine aguirreAún no hay calificaciones
- Fisiologia de AudicionDocumento62 páginasFisiologia de AudicionCaroline RibeiroAún no hay calificaciones
- Tema 8 FADocumento6 páginasTema 8 FA心夏老奶Aún no hay calificaciones
- Audicion (1-15)Documento3 páginasAudicion (1-15)leslie laraAún no hay calificaciones
- Tratado de Medicina Guyton Ty Hall-Cap 52 ResumenDocumento18 páginasTratado de Medicina Guyton Ty Hall-Cap 52 ResumenLudwing Pérez100% (2)
- El Sistema Auditivo y Su Influencia en La Adquisicion Del HablaDocumento8 páginasEl Sistema Auditivo y Su Influencia en La Adquisicion Del HablaPelisHD WeahkAún no hay calificaciones
- Audicion y EquilibrioDocumento16 páginasAudicion y EquilibrioAlexandra CuencaAún no hay calificaciones
- Tema 7Documento23 páginasTema 7Anabel GutierrezAún no hay calificaciones
- Sistema AuditivoDocumento4 páginasSistema AuditivoYesica ParraAún no hay calificaciones
- Free-T3.2 - La Audición-Gulag-FreeDocumento7 páginasFree-T3.2 - La Audición-Gulag-FreeCristina PereaAún no hay calificaciones
- El OidoDocumento12 páginasEl OidoYolanda CruzAún no hay calificaciones
- Tema 8 FADocumento7 páginasTema 8 FA心夏老奶Aún no hay calificaciones
- Resumen OidoDocumento5 páginasResumen OidoSilvina GrassoAún no hay calificaciones
- Fisiología Del OídoDocumento9 páginasFisiología Del OídoDoménica Romero VizuetaAún no hay calificaciones
- Guia-3C FISICA - El Oido HumanoDocumento4 páginasGuia-3C FISICA - El Oido Humanodragonrojo1964Aún no hay calificaciones
- AudiometriaDocumento24 páginasAudiometriaMary Nicole Aráuz AráuzAún no hay calificaciones
- Semana 10-3 y 11-1Documento5 páginasSemana 10-3 y 11-1Beatriz GarcíaAún no hay calificaciones
- Audición, Sentidos Somáticos y Sentidos QuímicosDocumento10 páginasAudición, Sentidos Somáticos y Sentidos QuímicosJoha RíosAún no hay calificaciones
- Tema 5 Psicoacústica y Percepción AuditivaDocumento83 páginasTema 5 Psicoacústica y Percepción AuditivaPau MarieAún no hay calificaciones
- Funcionamiento Del Oido HumanoDocumento29 páginasFuncionamiento Del Oido HumanoRuben PerezAún no hay calificaciones
- Sistema AuditivoDocumento22 páginasSistema AuditivoBruno OsorioAún no hay calificaciones
- Proceso Neurológico de La AudicionDocumento9 páginasProceso Neurológico de La AudicionCecilia UzateguiAún no hay calificaciones
- Física Acustica de La AudiciónDocumento7 páginasFísica Acustica de La Audicióndenisse.carvalhoAún no hay calificaciones
- Oidos y Audicion - CAP 10 - Equipo #3Documento17 páginasOidos y Audicion - CAP 10 - Equipo #3ANDRES DE JESUS PEREZ TORRESAún no hay calificaciones
- Oído InternoDocumento5 páginasOído InternoPedro CasanovaAún no hay calificaciones
- Sentido Del OídoDocumento5 páginasSentido Del OídoPablo GomezAún no hay calificaciones
- Tema 2 EL MECANISMO DE LA AUDICIÓNDocumento10 páginasTema 2 EL MECANISMO DE LA AUDICIÓNx3mgzAún no hay calificaciones
- Anatomía y Fisiología Del Oído.Documento6 páginasAnatomía y Fisiología Del Oído.Gianmauro Conforto MartinezAún no hay calificaciones
- Audiología IDocumento5 páginasAudiología IDavid Ivan Solis TrauquenAún no hay calificaciones
- Teoria de La AudicionDocumento11 páginasTeoria de La AudicionyajairaAún no hay calificaciones
- Extra 2. Sensibilidad AuditivaDocumento9 páginasExtra 2. Sensibilidad AuditivaAlbaAún no hay calificaciones
- Biofisica Del Sonido y AudicionDocumento76 páginasBiofisica Del Sonido y AudicionNatalia Brito NicasioAún no hay calificaciones
- Capítulo 53 Sentido de La AudiciónDocumento12 páginasCapítulo 53 Sentido de La AudiciónAlonso LunaAún no hay calificaciones
- Resumen de Sistema AuditivoDocumento2 páginasResumen de Sistema Auditivokarina Elizabeth Manzo SanchezAún no hay calificaciones
- El Sentido de La AudiciónDocumento13 páginasEl Sentido de La AudicióngabrielaAún no hay calificaciones
- Terapia Regenerativa en Hipoacusia NeurosensorialDocumento33 páginasTerapia Regenerativa en Hipoacusia NeurosensorialLina PérezAún no hay calificaciones
- Fisica Del SonidoDocumento5 páginasFisica Del SonidobbarbadillobAún no hay calificaciones
- Fisiología de La Audición.Documento67 páginasFisiología de La Audición.Cecilia Vitale SalasAún no hay calificaciones
- Sentidos EspecialesDocumento8 páginasSentidos EspecialesLeopoldo SerranoAún no hay calificaciones
- Biofísica de La AudiciónDocumento4 páginasBiofísica de La Audiciónyefry bossAún no hay calificaciones
- Sistema Auditivo PeriféricoDocumento11 páginasSistema Auditivo PeriféricoFabián RuizAún no hay calificaciones
- AudicionDocumento12 páginasAudicionJazmin LabbéAún no hay calificaciones
- Clase 1.3 - Fisiologia de La AudiconDocumento41 páginasClase 1.3 - Fisiologia de La AudiconmaurilioAún no hay calificaciones
- Conducto Auditivo ExternoDocumento5 páginasConducto Auditivo ExternoAbril.Aún no hay calificaciones
- AudicionDocumento15 páginasAudicionCamilo VeraAún no hay calificaciones
- El Mec. de La Percepción y Sus Caract. Cap 2Documento110 páginasEl Mec. de La Percepción y Sus Caract. Cap 2nano261191Aún no hay calificaciones
- Anatomía Del Sistema AuditivoDocumento12 páginasAnatomía Del Sistema AuditivoMarilú BelAún no hay calificaciones
- El OidoDocumento3 páginasEl Oidomauricio_lopez_gomez@hotmail.com80% (5)
- Anatomía y Fisiología de La AudiciónDocumento13 páginasAnatomía y Fisiología de La AudiciónAngela RodriguezAún no hay calificaciones
- Guia de Sentidos Especiales OidoDocumento7 páginasGuia de Sentidos Especiales OidoKarina Noemy Ramirez LarinAún no hay calificaciones
- Oido InternoDocumento55 páginasOido InternoCarito Pulache SiuAún no hay calificaciones
- Auditivo y VestibularDocumento12 páginasAuditivo y VestibularGenaro ValdovinosAún no hay calificaciones
- Componentes Del OídoDocumento6 páginasComponentes Del OídoJovanuzqa Jatsuky McAún no hay calificaciones
- Funciones y Partes Del OídoDocumento3 páginasFunciones y Partes Del OídobelenAún no hay calificaciones
- Lectura 5Documento9 páginasLectura 5bermejodalia1Aún no hay calificaciones
- Capítulo Vii. Grupo 2Documento65 páginasCapítulo Vii. Grupo 2salimabelvaleriodejimenezAún no hay calificaciones
- UntitledDocumento6 páginasUntitledcamilaAún no hay calificaciones
- UntitledDocumento13 páginasUntitledcamilaAún no hay calificaciones
- Test - Continuo Autista. Ángel RivièreDocumento1 páginaTest - Continuo Autista. Ángel RivièrecamilaAún no hay calificaciones
- 8-Point Penetration Aspiration Scale - Pas: Funcionamiento Categoría Puntaje Descripción de Los EventosDocumento1 página8-Point Penetration Aspiration Scale - Pas: Funcionamiento Categoría Puntaje Descripción de Los EventoscamilaAún no hay calificaciones
- Protocolos de Evaluación Del Adulto y Adulto Mayor: ManualdelfonoDocumento39 páginasProtocolos de Evaluación Del Adulto y Adulto Mayor: ManualdelfonocamilaAún no hay calificaciones
- El Sistema Vestibular y La CogniciónDocumento7 páginasEl Sistema Vestibular y La CognicióncamilaAún no hay calificaciones
- Neuroanatomía VestibularDocumento9 páginasNeuroanatomía VestibularcamilaAún no hay calificaciones
- Aplicación Clínica Del VHITDocumento9 páginasAplicación Clínica Del VHITcamilaAún no hay calificaciones
- Universidad Autónoma de Santo DomingoDocumento5 páginasUniversidad Autónoma de Santo DomingoJuan Francisco Baez PegueroAún no hay calificaciones
- Órgano de La Audición y El EquilibrioDocumento23 páginasÓrgano de La Audición y El EquilibrioEstëfany Isä SötoAún no hay calificaciones
- Apunte de OtoDocumento33 páginasApunte de OtoRicardo Ramos AndradeAún no hay calificaciones
- Proyecto Integrador PDFDocumento23 páginasProyecto Integrador PDFValentinaAún no hay calificaciones
- Copia de Expo Otorrino TeoriaDocumento22 páginasCopia de Expo Otorrino TeoriaDANIELA NICOLE MAINGON SEVILLAAún no hay calificaciones
- OtosDocumento14 páginasOtosJesus Avila100% (1)
- T Uce 0020 Cdi 480Documento42 páginasT Uce 0020 Cdi 480Pablo GarcíaAún no hay calificaciones
- Bases LogoaudiometríaDocumento18 páginasBases LogoaudiometríaLourdes C. Triviño GómezAún no hay calificaciones
- Anatomía Del OidoDocumento2 páginasAnatomía Del OidoAlexandra castroAún no hay calificaciones
- ColesteatomaDocumento4 páginasColesteatomaluzAún no hay calificaciones
- AnatomÍa y FisiologÍa Del OídoDocumento48 páginasAnatomÍa y FisiologÍa Del Oídodomingoam31170188% (8)
- Album Patologías de OídoDocumento21 páginasAlbum Patologías de OídoyeniAún no hay calificaciones
- Sistema Vestibular OidoDocumento11 páginasSistema Vestibular OidoAnonymous WKtWv4oAún no hay calificaciones
- Presentacion Internado Carlos Van BurenDocumento32 páginasPresentacion Internado Carlos Van BurenNataly FernandaAún no hay calificaciones
- Microfonicas CoclearesDocumento4 páginasMicrofonicas CoclearesNoe CausaranoAún no hay calificaciones
- Clase 1 ZDocumento55 páginasClase 1 ZValeria PradoAún no hay calificaciones
- Terapia Ocupacional en Personas Con Déficits AuditivosDocumento22 páginasTerapia Ocupacional en Personas Con Déficits AuditivosCamila NoriegaAún no hay calificaciones
- Partes Del Oído y Partes de La OrejaDocumento3 páginasPartes Del Oído y Partes de La OrejaEve ColomboAún no hay calificaciones
- Rotulado OidoDocumento32 páginasRotulado OidoNicoleFuentesGalleguillosAún no hay calificaciones
- Mapa Conceptual - Oído InternoDocumento1 páginaMapa Conceptual - Oído InternoLucas PicadaAún no hay calificaciones
- Ayuda PruebaDocumento3 páginasAyuda PruebaveronicaAún no hay calificaciones
- El OidoDocumento6 páginasEl Oidomojica coronado FLAún no hay calificaciones
- MiringoplastiaDocumento42 páginasMiringoplastiaGuillermo Stipech100% (1)
- Gemo-005 Guia Tecnica AudiometriaDocumento20 páginasGemo-005 Guia Tecnica AudiometriaW Andres Jajoy Juajibioy100% (1)
- OtotoxicidadDocumento7 páginasOtotoxicidadClaudia TorresAún no hay calificaciones
- Fisiología de La AudiciónDocumento13 páginasFisiología de La AudicióncamilaAún no hay calificaciones