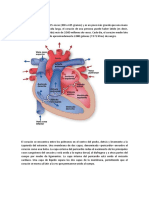
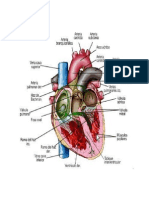
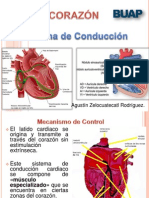
También podría gustarte
- Fisiologia CardiacaDocumento7 páginasFisiologia CardiacaMercedes CanalesAún no hay calificaciones
- Manual Usuario LightSheerDocumento28 páginasManual Usuario LightSheerjoseaselopez100% (2)
- Capacitacion Nom 009 Stps 2011Documento37 páginasCapacitacion Nom 009 Stps 2011LUIS ALBERTO VENEGAS90% (10)
- Tema 3.PROPIEDADES ELECTROFISIOLÓGICAS DEL MIOCARDIO CONTRÁCTIL Y ESPECIALIZADODocumento7 páginasTema 3.PROPIEDADES ELECTROFISIOLÓGICAS DEL MIOCARDIO CONTRÁCTIL Y ESPECIALIZADOyuri goitiaAún no hay calificaciones
- Fisiología Cardíaca: Sistema de ConducciónDocumento36 páginasFisiología Cardíaca: Sistema de ConducciónCarlos AvilaAún no hay calificaciones
- TEMA 6_SISTEMA CARDIOVASCULAR.pdf APUNTESDocumento34 páginasTEMA 6_SISTEMA CARDIOVASCULAR.pdf APUNTESCode - NoSoyjessAún no hay calificaciones
- ANATOMÍA MICROSCÓPICA Músculo Cardíaco El Miocardio o Músculo Cardíaco Está Formado Por Fibras Musculares Estriadas Más Cortas y Menos Circulares Que Las Fibras Del Músculo EsqueléticoDocumento4 páginasANATOMÍA MICROSCÓPICA Músculo Cardíaco El Miocardio o Músculo Cardíaco Está Formado Por Fibras Musculares Estriadas Más Cortas y Menos Circulares Que Las Fibras Del Músculo EsqueléticoEstherAún no hay calificaciones
- Autor Anónimo - EKG Con Buenas ImágenesDocumento12 páginasAutor Anónimo - EKG Con Buenas ImágenesJuan Carlos AlfaroAún no hay calificaciones
- Fisiologias Cardiaca PDFDocumento7 páginasFisiologias Cardiaca PDFRonny Chilon TroncosAún no hay calificaciones
- Sistema Cardiovascular 2 - JEsús GonzálezDocumento13 páginasSistema Cardiovascular 2 - JEsús GonzálezJesús GonzálezAún no hay calificaciones
- Estimulacion Ritmica y Conductividad CardiacaDocumento5 páginasEstimulacion Ritmica y Conductividad CardiacaJenniffer EspinalesAún no hay calificaciones
- Actividad Eléctrica Del CorazónDocumento18 páginasActividad Eléctrica Del CorazónAldo SanabriaAún no hay calificaciones
- INTRODUCCION A LA FISIOLOGIA CARDIACADocumento16 páginasINTRODUCCION A LA FISIOLOGIA CARDIACAMartin PiattiAún no hay calificaciones
- CorazonDocumento19 páginasCorazonDark-Win M. ChalkoAún no hay calificaciones
- El Músculo CardíacoDocumento2 páginasEl Músculo CardíacoEDUARDO MARQUEZ DOMINGUEZAún no hay calificaciones
- Guía del Sistema CardiovascularDocumento10 páginasGuía del Sistema CardiovascularBárbara AguileraAún no hay calificaciones
- El Sistema de Conducción o Nodal Del CorazónDocumento7 páginasEl Sistema de Conducción o Nodal Del CorazónjhonnyAún no hay calificaciones
- Funcionamiento y Estructura Del CorazónDocumento3 páginasFuncionamiento y Estructura Del CorazónMaria JoseAún no hay calificaciones
- SarcomeroDocumento7 páginasSarcomeroIliana NarváezAún no hay calificaciones
- Cardiovascular I - Conceptos introductoriosDocumento13 páginasCardiovascular I - Conceptos introductoriosNatalia Oscco VenturaAún no hay calificaciones
- 4 CardiovascularDocumento8 páginas4 CardiovascularmariarofalconAún no hay calificaciones
- POTENCIAL DE MEMBRANA Y CARDIOVASCULADocumento7 páginasPOTENCIAL DE MEMBRANA Y CARDIOVASCULAJacki ServianAún no hay calificaciones
- TP 15,16 y 17 Fisiología de Sistema Cardiovascular I y IIDocumento24 páginasTP 15,16 y 17 Fisiología de Sistema Cardiovascular I y IIGabriel SAún no hay calificaciones
- Tema Fisiologia CardiacaDocumento6 páginasTema Fisiologia CardiacaDalia JimenezAún no hay calificaciones
- 1era Semana CardioDocumento37 páginas1era Semana CardioFlorencia GuastalliAún no hay calificaciones
- Aparato Cardiovascular 1T1Documento41 páginasAparato Cardiovascular 1T1Marycielo LmAún no hay calificaciones
- Electrocardiograma FinalDocumento23 páginasElectrocardiograma FinalLaura SanabriaAún no hay calificaciones
- Activación Eléctrica Del CorazónDocumento3 páginasActivación Eléctrica Del CorazónGuisela Paredes AlaniaAún no hay calificaciones
- Sistema Electrico Del CorazónDocumento3 páginasSistema Electrico Del CorazónFlor MartinezAún no hay calificaciones
- Sitema CardiovascularDocumento13 páginasSitema CardiovascularRo AvilaAún no hay calificaciones
- SistemaCardiovascularContraccionMuscularDocumento12 páginasSistemaCardiovascularContraccionMuscularSHEYLA DEL VICTORIA GUILLENAún no hay calificaciones
- Fbbva Librocorazon Cap3Documento8 páginasFbbva Librocorazon Cap3CristhianEsequielQuispelayoRamosAún no hay calificaciones
- Función cardíaca y válvulasDocumento6 páginasFunción cardíaca y válvulasAstrid LainezAún no hay calificaciones
- El Corazón Como BombaDocumento10 páginasEl Corazón Como BombaReina PerezAún no hay calificaciones
- ARRITMIASDocumento21 páginasARRITMIASJoselyn RodriguezAún no hay calificaciones
- Funcion CardiacaDocumento51 páginasFuncion CardiacaLeonel Sanchez GarciaAún no hay calificaciones
- Latido cardíaco y ciclo cardíacoDocumento9 páginasLatido cardíaco y ciclo cardíacoRamon Andres Gonzalez SayleAún no hay calificaciones
- Expo de FisiologiaDocumento3 páginasExpo de FisiologiaCarolina Gonzales NuñesAún no hay calificaciones
- Sistema Exito ConductorDocumento10 páginasSistema Exito ConductorZadith Milagros Tello Gonzales100% (1)
- Cuestionario de Músculo CardiacoDocumento3 páginasCuestionario de Músculo CardiacoJesús Leyva GarcíaAún no hay calificaciones
- Fisiologia CardiacaDocumento15 páginasFisiologia CardiacaAgnódice de AtenasAún no hay calificaciones
- Excitoconductor y EKGDocumento16 páginasExcitoconductor y EKGAlbert FigueredoAún no hay calificaciones
- Fisiología Cardíaca-MedGeneralIIDocumento8 páginasFisiología Cardíaca-MedGeneralIIAndrea PaosulineAún no hay calificaciones
- Musculo Cardiaco Preparatoria EvaDocumento15 páginasMusculo Cardiaco Preparatoria Evanachocoolpro855Aún no hay calificaciones
- Bloque 2 CardiorrespiratorioDocumento29 páginasBloque 2 CardiorrespiratorioJosé EireneAún no hay calificaciones
- Fisiologia CardiacaDocumento19 páginasFisiologia CardiacaRosa BlancoAún no hay calificaciones
- CORAZONEXPODocumento17 páginasCORAZONEXPOAgus ZelRodAún no hay calificaciones
- CORAZONDocumento30 páginasCORAZONXimena FloresAún no hay calificaciones
- TP Sistema CirculatorioDocumento3 páginasTP Sistema CirculatorioGerónimo SikAún no hay calificaciones
- Sistema de Conducción Eléctrica Del CorazónDocumento37 páginasSistema de Conducción Eléctrica Del CorazónMagy Cruz MarAún no hay calificaciones
- Cardio 1 PDFDocumento37 páginasCardio 1 PDFEuge OrihuelaAún no hay calificaciones
- Anatomía Del Sistema de ConducciónDocumento5 páginasAnatomía Del Sistema de ConducciónDaniela IslasAún no hay calificaciones
- Ligaduras de StaDocumento11 páginasLigaduras de StaSELENE VANESSA CHOZO MESTANZAAún no hay calificaciones
- Anatomía Del CorazónDocumento65 páginasAnatomía Del CorazónLiliana Beatriz MendezAún no hay calificaciones
- SISTEMA DE CONDUCCIÓN ELECTRICA CARDIACADocumento5 páginasSISTEMA DE CONDUCCIÓN ELECTRICA CARDIACAkenia adhinary renteria cabreraAún no hay calificaciones
- Apuntes Del Sistema CardiovascularDocumento23 páginasApuntes Del Sistema CardiovascularAdriana López-ValverdeAún no hay calificaciones
- Músculo cardíaco: estructura y función de las células miocárdicasDocumento12 páginasMúsculo cardíaco: estructura y función de las células miocárdicasZuleyma SánchezAún no hay calificaciones
- Anatomia y Fisiologia Del Sistema de Conducción (Modulo 1)Documento7 páginasAnatomia y Fisiologia Del Sistema de Conducción (Modulo 1)Patty LazcanoAún no hay calificaciones
- Mejorar la Circulación Sanguínea de Forma Natural: Sanar el Sistema Circulatorio y inmunológico para Prevenir Enfermedades, Aumentar la Energía, Aliviar el Estrés y Sentirte Mejor que NuncaDe EverandMejorar la Circulación Sanguínea de Forma Natural: Sanar el Sistema Circulatorio y inmunológico para Prevenir Enfermedades, Aumentar la Energía, Aliviar el Estrés y Sentirte Mejor que NuncaCalificación: 4.5 de 5 estrellas4.5/5 (7)
- Introducción Básica a la Microcirculación de la Sangre: La Siguiente Generación en Cuidados de la SaludDe EverandIntroducción Básica a la Microcirculación de la Sangre: La Siguiente Generación en Cuidados de la SaludAún no hay calificaciones
- Semana 5 Cultura e Identidad CiudadanaDocumento33 páginasSemana 5 Cultura e Identidad CiudadanaJose Luis BustamanteAún no hay calificaciones
- Semana 2 Taxonomía y Nomenclatura - Replicación ViralDocumento36 páginasSemana 2 Taxonomía y Nomenclatura - Replicación ViralJose Luis BustamanteAún no hay calificaciones
- Clase 7 Elisa SNDocumento21 páginasClase 7 Elisa SNAlyssa Li SalazarAún no hay calificaciones
- Clase 3 Patogenia ViralDocumento43 páginasClase 3 Patogenia ViralAlyssa Li SalazarAún no hay calificaciones
- Desarrollo Humano.Documento17 páginasDesarrollo Humano.Jose Luis BustamanteAún no hay calificaciones
- Semana 2 Periodización de La Historia Del PerúDocumento34 páginasSemana 2 Periodización de La Historia Del PerúJose Luis BustamanteAún no hay calificaciones
- Semana 4 Guerra Del Pacífico.Documento39 páginasSemana 4 Guerra Del Pacífico.Jose Luis BustamanteAún no hay calificaciones
- Rabia Canina - VirologiaDocumento9 páginasRabia Canina - VirologiaJose Luis BustamanteAún no hay calificaciones
- Semana 3 Independencia Del Perú.Documento27 páginasSemana 3 Independencia Del Perú.Jose Luis BustamanteAún no hay calificaciones
- Semana 7 Práctica de Fisiología de Grupos SanguíneosDocumento8 páginasSemana 7 Práctica de Fisiología de Grupos SanguíneosJose Luis BustamanteAún no hay calificaciones
- Realidad Nacional VeterinariaDocumento24 páginasRealidad Nacional VeterinariaJose Luis BustamanteAún no hay calificaciones
- Semana 4 Teoria Fisiologia Del Sistema Nervioso CentralDocumento15 páginasSemana 4 Teoria Fisiologia Del Sistema Nervioso CentralJose Luis BustamanteAún no hay calificaciones
- Semana 6 Práctica Fisiología SanguíneaDocumento13 páginasSemana 6 Práctica Fisiología SanguíneaJose Luis BustamanteAún no hay calificaciones
- PDF 2022 II Clases Teorica Generalidades de La NutricionDocumento55 páginasPDF 2022 II Clases Teorica Generalidades de La NutricionJose Luis BustamanteAún no hay calificaciones
- Clases Teoricas Metabolismo de Los CarbohidratosDocumento42 páginasClases Teoricas Metabolismo de Los CarbohidratosJose Luis BustamanteAún no hay calificaciones
- Detección de antígenos virales mediante ELISADocumento4 páginasDetección de antígenos virales mediante ELISAJose Luis BustamanteAún no hay calificaciones
- Casos Clinicos Orina (Clase 3) UpsjbDocumento58 páginasCasos Clinicos Orina (Clase 3) UpsjbJose Luis BustamanteAún no hay calificaciones
- Características Fenotípicas de La Fibra, Vellón en La Captura (Chaku) y Grupos Sociales de La Vicuña.Documento9 páginasCaracterísticas Fenotípicas de La Fibra, Vellón en La Captura (Chaku) y Grupos Sociales de La Vicuña.Jose Luis BustamanteAún no hay calificaciones
- boldmdocumentsDialogoMedicoPacienteQuechua PDFDocumento96 páginasboldmdocumentsDialogoMedicoPacienteQuechua PDFJose Luis BustamanteAún no hay calificaciones
- Raza YorkshireDocumento3 páginasRaza YorkshireJose Luis BustamanteAún no hay calificaciones
- Informe4 Patos y Pavo ZootecniaDocumento8 páginasInforme4 Patos y Pavo ZootecniaJose Luis BustamanteAún no hay calificaciones
- Descriptores de CuyesDocumento13 páginasDescriptores de CuyesJose Luis BustamanteAún no hay calificaciones
- Fisiologia de La PreñezDocumento4 páginasFisiologia de La PreñezJose Luis BustamanteAún no hay calificaciones
- Identificar Las Características Raciales Del Caballo Peruano de Paso.Documento8 páginasIdentificar Las Características Raciales Del Caballo Peruano de Paso.Jose Luis BustamanteAún no hay calificaciones
- Plan+de+Cuidados+Al+Paciente+Apendicectomizado 2010Documento16 páginasPlan+de+Cuidados+Al+Paciente+Apendicectomizado 2010Ale Boglio CAún no hay calificaciones
- FORMATO Administrar InmunobiolDocumento4 páginasFORMATO Administrar InmunobiolFernando CastañedaAún no hay calificaciones
- Miasis Buco-Maxilo-Facial Reporte de Un CasoDocumento6 páginasMiasis Buco-Maxilo-Facial Reporte de Un CasoVirtualdental MexicoAún no hay calificaciones
- Alerta Digemid #08-2023Documento2 páginasAlerta Digemid #08-2023Mirian Ruth Yucra CoylaAún no hay calificaciones
- La Noche Boca ArribaDocumento6 páginasLa Noche Boca ArribaRodrigo Andrés Valenzuela Zura100% (1)
- Asimetría CerebralDocumento43 páginasAsimetría CerebralJINETH PAOLA RIVERA CASTROAún no hay calificaciones
- Nuevo Formato Psicotropicos 2022Documento1 páginaNuevo Formato Psicotropicos 2022EdwinRivasAún no hay calificaciones
- Alelos MúltiplesDocumento5 páginasAlelos MúltiplesMalicia VázquezAún no hay calificaciones
- La TosDocumento17 páginasLa TosRosaura Gutierrez QuispeAún no hay calificaciones
- Reporte de FarmacodinamiaDocumento40 páginasReporte de FarmacodinamiaCinthya Castillo100% (1)
- Examen Mensual de PL RVDocumento2 páginasExamen Mensual de PL RVAnonymous lEcCLNuEAún no hay calificaciones
- DolorDocumento1 páginaDolorLaura BribiescaAún no hay calificaciones
- Ix Ciclo - Grupo A Caso Clinico - Dra MezaDocumento5 páginasIx Ciclo - Grupo A Caso Clinico - Dra MezaCARLOS SALVADOR GERONIMO ESPINOAún no hay calificaciones
- PPP Nutrición y Estilos VidaDocumento2 páginasPPP Nutrición y Estilos VidanelsonjmbAún no hay calificaciones
- Preguntas y Respuestas TabaquismoDocumento4 páginasPreguntas y Respuestas TabaquismoMary LuzAún no hay calificaciones
- End 3Documento4 páginasEnd 3Luis ColomaAún no hay calificaciones
- FeocromocitomaDocumento9 páginasFeocromocitomaCh LaiAún no hay calificaciones
- David Lopez Capape Lesiones Articulacion LisfrancDocumento7 páginasDavid Lopez Capape Lesiones Articulacion Lisfrancljaramillo_25Aún no hay calificaciones
- CN - El Plato Del Bien ComerDocumento3 páginasCN - El Plato Del Bien ComerNepomuceno Camila JaraAún no hay calificaciones
- Vitaminoterapia 2Documento9 páginasVitaminoterapia 2badylocoAún no hay calificaciones
- Tesen 0Documento2 páginasTesen 0MARA PEREZ CASTROAún no hay calificaciones
- Aspiracion de SecrecionesDocumento1 páginaAspiracion de SecrecionesOlivia Fernanda Verduzco CastroAún no hay calificaciones
- Pediatría PreventivaDocumento24 páginasPediatría PreventivaMishell PalaciosAún no hay calificaciones
- Infografía, Enfermera Auxiliar Del Médico..Documento1 páginaInfografía, Enfermera Auxiliar Del Médico..Mauricio RodriguezAún no hay calificaciones
- Programa de Capacitación - 606Documento29 páginasPrograma de Capacitación - 606Michelle CotaAún no hay calificaciones
- ESAVIDocumento61 páginasESAVITaliza Alberca EspinoAún no hay calificaciones
- Gerencia en SaludDocumento10 páginasGerencia en SaludSabrina YalliAún no hay calificaciones
- MATRIZDocumento5 páginasMATRIZMaría Alejandra López AzaAún no hay calificaciones