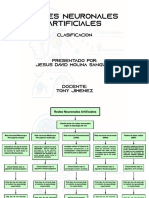
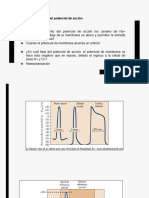
También podría gustarte
- Cómo se comunican las neuronas: El milagro de la transmisión sinápticaDe EverandCómo se comunican las neuronas: El milagro de la transmisión sinápticaAún no hay calificaciones
- Articulo Proyecto Prótesis Con Piel Artificial Con Nanoparticulas Dr. Antonio Faustino Muñoz y Otr.-1Documento9 páginasArticulo Proyecto Prótesis Con Piel Artificial Con Nanoparticulas Dr. Antonio Faustino Muñoz y Otr.-1Sebastián MacíasAún no hay calificaciones
- Tecnicas de NeuroimagenDocumento53 páginasTecnicas de NeuroimagenMaria Jose SimonAún no hay calificaciones
- 100 circuitos de shields para arduino (español)De Everand100 circuitos de shields para arduino (español)Calificación: 3.5 de 5 estrellas3.5/5 (3)
- Paper EogDocumento4 páginasPaper EogGonzalo BordaAún no hay calificaciones
- Ia Predicción de Variables en Planta ConcentradoraDocumento22 páginasIa Predicción de Variables en Planta ConcentradoraMICHAEL CUETO MERCADOAún no hay calificaciones
- Lab Bioinstru3corte1 Abcdpdf PDF A WordDocumento6 páginasLab Bioinstru3corte1 Abcdpdf PDF A WordCRISTIAN LEONARDO SUAREZ GARCIAAún no hay calificaciones
- Sistema de Monitoreo de Las Ondas Cerebrales (ElectroencefalogramaDocumento12 páginasSistema de Monitoreo de Las Ondas Cerebrales (ElectroencefalogramaBeatrizBenitezAún no hay calificaciones
- Lab Bioinstru3corte1Documento6 páginasLab Bioinstru3corte1CRISTIAN LEONARDO SUAREZ GARCIAAún no hay calificaciones
- Informe 9Documento5 páginasInforme 9Brayan Stiven Torres NincoAún no hay calificaciones
- Diciembre 2012Documento6 páginasDiciembre 2012Belén BrownAún no hay calificaciones
- Diciembre 2012-Fusionado-Fusionado-FusionadoDocumento115 páginasDiciembre 2012-Fusionado-Fusionado-FusionadoEvangelina Ruiz100% (1)
- Técnicas y Métodos Electrofisiológicos para El Estudio de La Relación Entre Cerebro y Conducta.Documento7 páginasTécnicas y Métodos Electrofisiológicos para El Estudio de La Relación Entre Cerebro y Conducta.Wendy Marleen Rodriguez GutierrezAún no hay calificaciones
- Sinapsis IDocumento58 páginasSinapsis IEvelyn RodriguezAún no hay calificaciones
- Técnicas de NeuroimagenDocumento11 páginasTécnicas de NeuroimagenRodrigo Solar Navarrete100% (1)
- Actividad No 4 Mapa Conceptual Redes Neuronales ArtificialesDocumento3 páginasActividad No 4 Mapa Conceptual Redes Neuronales Artificialesjesus100% (1)
- 3 Tema-MagnetoDocumento13 páginas3 Tema-MagnetocarlotaAún no hay calificaciones
- Campos Electromagnéticos Neuronales - ¿Sirven para Algo - Antroporama - Antroporama PDFDocumento6 páginasCampos Electromagnéticos Neuronales - ¿Sirven para Algo - Antroporama - Antroporama PDFSebastian SolanoAún no hay calificaciones
- Resumen Redes NeuronalesDocumento7 páginasResumen Redes NeuronalesGerman E Ruiz CorralesAún no hay calificaciones
- Investigacion: Neocognitron. Redes PCADocumento13 páginasInvestigacion: Neocognitron. Redes PCAConstance Villacreses0% (1)
- PWM AnaDocumento10 páginasPWM Anaarmando herrera tellezAún no hay calificaciones
- Morfología Macroscópica Funcional Lissette Gomes (Prof. Anny Vasquez)Documento7 páginasMorfología Macroscópica Funcional Lissette Gomes (Prof. Anny Vasquez)Lissette Gomes AlbarranAún no hay calificaciones
- Mapa Conceptual Redes NeuronalesDocumento3 páginasMapa Conceptual Redes NeuronalesWILMER ANDRES CASTILLA CAROAún no hay calificaciones
- Proyecto Analisis de Senales y Sistemas Iza Martinez Puchuri y VidalDocumento5 páginasProyecto Analisis de Senales y Sistemas Iza Martinez Puchuri y VidalSköulls SpartAún no hay calificaciones
- Paper Eog Copa Cruz MamaniDocumento5 páginasPaper Eog Copa Cruz Mamaniadrian ramos ramosAún no hay calificaciones
- Efalografía Convencional en Pediatría:: ElectroencDocumento4 páginasEfalografía Convencional en Pediatría:: ElectroencsusanaAún no hay calificaciones
- Trabajo de Investigación RNADocumento11 páginasTrabajo de Investigación RNAJose SilvaAún no hay calificaciones
- Tercera Entrega Electromiograma - Gomez, Martinez, Osorio & RodriguezDocumento17 páginasTercera Entrega Electromiograma - Gomez, Martinez, Osorio & RodriguezNicolas OrozcoAún no hay calificaciones
- Rehabilitacion Z, P, CDocumento6 páginasRehabilitacion Z, P, CLeonardo Fabio Velásquez IncaAún no hay calificaciones
- RN Sesion5 B Apli Alarmas ScadaDocumento17 páginasRN Sesion5 B Apli Alarmas ScadaKarin CMAún no hay calificaciones
- Ip Labo3 DW PDFDocumento4 páginasIp Labo3 DW PDFwilsonAún no hay calificaciones
- Arch IvoDocumento18 páginasArch IvoRonny YupanquiAún no hay calificaciones
- Introduccion BciDocumento55 páginasIntroduccion BciJavier Ferney Castillo GarciaAún no hay calificaciones
- 50 PDFDocumento5 páginas50 PDFfrank naranjoAún no hay calificaciones
- Actividad 1.2 - Procesamiento Digital de Señales ElectroencefalográficasDocumento22 páginasActividad 1.2 - Procesamiento Digital de Señales ElectroencefalográficasDiegoIramLunaCastañedaAún no hay calificaciones
- Informe1 - Introduccion A La Computacion NeuronalDocumento15 páginasInforme1 - Introduccion A La Computacion NeuronalCarlos GuarinAún no hay calificaciones
- Cap 10Documento38 páginasCap 10Francisco CermeñoAún no hay calificaciones
- Neurociencia. Introducción de Las Matemáticas Aplicadas en NeurocienciaDocumento7 páginasNeurociencia. Introducción de Las Matemáticas Aplicadas en NeurocienciaJoel EduardoAún no hay calificaciones
- Paper Eog Copa Cruz MamaniDocumento5 páginasPaper Eog Copa Cruz Mamaniadrian ramos ramosAún no hay calificaciones
- Implantes Cocleares y de TroncoDocumento20 páginasImplantes Cocleares y de TroncoSchitton Gonzalez100% (1)
- Ses.9 - Tecnicas de NeuroimagenDocumento15 páginasSes.9 - Tecnicas de NeuroimagenSarita Sujey Diaz SalvatierraAún no hay calificaciones
- Proyecto Experimental FinalDocumento9 páginasProyecto Experimental Finalannetg2204Aún no hay calificaciones
- SINAPSISDocumento5 páginasSINAPSISJireh TorresAún no hay calificaciones
- Taller RNADocumento20 páginasTaller RNAAlberto Jimenez MiguelAún no hay calificaciones
- UPC 610 AGUI 2008 2404 Neurocie LDocumento72 páginasUPC 610 AGUI 2008 2404 Neurocie LIngrid MendozaAún no hay calificaciones
- Distintos Métodos de Investigación deDocumento2 páginasDistintos Métodos de Investigación deMariela FloresAún no hay calificaciones
- Impulso Nervioso, Sinapsis y NeurotransmisoresDocumento17 páginasImpulso Nervioso, Sinapsis y NeurotransmisoresDagoberto Andres Fuentes Solis100% (2)
- Cuadro ComparativoDocumento3 páginasCuadro ComparativoHerald Leon MelgarAún no hay calificaciones
- Poster MultiDocumento3 páginasPoster MultiGU JAAún no hay calificaciones
- PR Ctica3 Lab Contempo1Documento4 páginasPR Ctica3 Lab Contempo1Sandi HernándezAún no hay calificaciones
- Metodos de Exploracion Neuroimagen PDFDocumento51 páginasMetodos de Exploracion Neuroimagen PDFCecilia Paulina Concha Gonzalez.100% (2)
- Revista Cubana de Ciencias Informáticas 1994-1536: Issn: Rcci@uci - CuDocumento9 páginasRevista Cubana de Ciencias Informáticas 1994-1536: Issn: Rcci@uci - CuVEDER MIGUEL TORRES BENITESAún no hay calificaciones
- NEURONASDocumento5 páginasNEURONASHaiver Neftali Hernández CastroAún no hay calificaciones
- BIO021 S3 Proteína GDocumento100 páginasBIO021 S3 Proteína GROCIO SINAI HERNANDEZ JUAREZAún no hay calificaciones
- BIO021 + AcetilcolinaDocumento129 páginasBIO021 + AcetilcolinaROCIO SINAI HERNANDEZ JUAREZAún no hay calificaciones
- Informe 10Documento6 páginasInforme 10Luisa Fernanda Ramirez VelascoAún no hay calificaciones
- Practica N 5Documento5 páginasPractica N 5Felo OnofaAún no hay calificaciones
- Telemetría de Eventos SísmicoDocumento16 páginasTelemetría de Eventos SísmicoCarlos Alberto Henao BaenaAún no hay calificaciones
- Tecnicas de Neuroimagen y SensopercepcionDocumento40 páginasTecnicas de Neuroimagen y SensopercepcionLeonardo MiñoAún no hay calificaciones
- YinChen Electrogastrography - En.esDocumento13 páginasYinChen Electrogastrography - En.esAlexander Morales BernalAún no hay calificaciones
- OptogeneticssDocumento13 páginasOptogeneticssAlexander Morales BernalAún no hay calificaciones
- Modulation of H-Reflex Responses and Frequency-DependentDocumento15 páginasModulation of H-Reflex Responses and Frequency-DependentAlexander Morales BernalAún no hay calificaciones
- Optogenética Un Panorama General, Histórico y Revisión Bibliográfica de Aplicaciones en Enfermedades NeurodegenerativasDocumento20 páginasOptogenética Un Panorama General, Histórico y Revisión Bibliográfica de Aplicaciones en Enfermedades NeurodegenerativasAlexander Morales BernalAún no hay calificaciones
- Elsistemainternacionaldeunidades WebDocumento113 páginasElsistemainternacionaldeunidades WebJulio PinedaAún no hay calificaciones
- Pettinelli Natalia TD 2020Documento175 páginasPettinelli Natalia TD 2020Alexander Morales BernalAún no hay calificaciones
- Grafo 3Documento75 páginasGrafo 3Jaime Adolfo Mejia LeónAún no hay calificaciones
- Evaluación de La Incertidumbre en DatosDocumento43 páginasEvaluación de La Incertidumbre en Datosikki_kueilanAún no hay calificaciones
- Grafo 3Documento75 páginasGrafo 3Jaime Adolfo Mejia LeónAún no hay calificaciones
- (Tac) Spinelli CarlaDocumento55 páginas(Tac) Spinelli CarlaTeresita RSAún no hay calificaciones
- 3.2 Propagacion de ErroresDocumento14 páginas3.2 Propagacion de Errores63rAún no hay calificaciones
- Solucionario Fisica de Resnick - Halliday - Krane - 5ta Ed Vol 1Documento315 páginasSolucionario Fisica de Resnick - Halliday - Krane - 5ta Ed Vol 1Alan Francis100% (1)
- Problemario de TermodinamicaDocumento3 páginasProblemario de TermodinamicaVianey FloresAún no hay calificaciones
- Los Procesos Geológicos Externos y El RelieveDocumento54 páginasLos Procesos Geológicos Externos y El RelieveAaron CastroAún no hay calificaciones
- 2º Examen Parcial de Electroquímica Respuestas PDFDocumento4 páginas2º Examen Parcial de Electroquímica Respuestas PDFMarlene Karina Salinas QuinteroAún no hay calificaciones
- Curso de Masaje 1ra. ParteDocumento25 páginasCurso de Masaje 1ra. ParteVirginia MendezAún no hay calificaciones
- Proroot Arroz Finca El Progreso 2013 PDFDocumento45 páginasProroot Arroz Finca El Progreso 2013 PDFcesarAún no hay calificaciones
- Introducción Al Derecho Internacional HumanitarioDocumento37 páginasIntroducción Al Derecho Internacional HumanitarioJordanRoldánAún no hay calificaciones
- Arreglo de CamasDocumento34 páginasArreglo de Camasdenisse castro100% (1)
- Edan OgboniDocumento5 páginasEdan OgboniAwosade OlaifaAún no hay calificaciones
- Teoria Del Apego BowlbyDocumento23 páginasTeoria Del Apego Bowlbymgsssgm100% (2)
- Matriz de Identificación de Peligros TRANSPORTES JAACS Soprole ConducciónDocumento2 páginasMatriz de Identificación de Peligros TRANSPORTES JAACS Soprole ConducciónSebastian MurphyAún no hay calificaciones
- 5 Poniendo Atención Al Macho Reproductor PDFDocumento4 páginas5 Poniendo Atención Al Macho Reproductor PDFavexperuAún no hay calificaciones
- Turismo de Salud Islas CanariasDocumento64 páginasTurismo de Salud Islas CanariasDaiana MAún no hay calificaciones
- Ensayo de JarrasDocumento5 páginasEnsayo de JarrasDavidPiravanAún no hay calificaciones
- Actividad de Aprendizaje 16 Evidencia 5Documento15 páginasActividad de Aprendizaje 16 Evidencia 5Mery SuarezAún no hay calificaciones
- Equipamiento LaboratorioDocumento8 páginasEquipamiento Laboratorioisidro barrientosAún no hay calificaciones
- Tarea#5 Lesby DiazDocumento3 páginasTarea#5 Lesby DiazMichael PettoAún no hay calificaciones
- C15. Disfunción Neuromuscular en La SepsisDocumento25 páginasC15. Disfunción Neuromuscular en La SepsisCristóbal Miño MoralesAún no hay calificaciones
- Teoría de La Identidad Erik Erikson InfografiaDocumento2 páginasTeoría de La Identidad Erik Erikson Infografiamariana.solis138Aún no hay calificaciones
- Ficha Antropometrica Sena PDFDocumento2 páginasFicha Antropometrica Sena PDFMelisa LondoñoAún no hay calificaciones
- Ficha Tecnica TRIHAL 3150KVA - 25000V - 420V - ECODISEÑODocumento1 páginaFicha Tecnica TRIHAL 3150KVA - 25000V - 420V - ECODISEÑOMarioyfernanda Guerra MuruaAún no hay calificaciones
- Plan-Manos A La siembra-FELIXALVARADODocumento11 páginasPlan-Manos A La siembra-FELIXALVARADOFellixRafaelAlvarado100% (1)
- Ensayo Mercado Meta BAYERDocumento5 páginasEnsayo Mercado Meta BAYERCarylAún no hay calificaciones
- Proyectos Etwinning de FP 14 IdeasDocumento38 páginasProyectos Etwinning de FP 14 IdeasLuis C. Peinado BravoAún no hay calificaciones
- Planeacion Docente Institucional Membrete.Documento2 páginasPlaneacion Docente Institucional Membrete.jhonAún no hay calificaciones
- Realidad Aumentada en La MedicinaDocumento8 páginasRealidad Aumentada en La MedicinaValeriaAún no hay calificaciones
- Diferencial Semantico - Actitudes Al PsicologoDocumento2 páginasDiferencial Semantico - Actitudes Al PsicologoJonathan PérezAún no hay calificaciones
- FPIPS-102 Vision Del Negocio-1Documento11 páginasFPIPS-102 Vision Del Negocio-1diegx217Aún no hay calificaciones
- GERMICIDA BASE DE AMONIO CUATERNARIO 5a Generación.v4Documento10 páginasGERMICIDA BASE DE AMONIO CUATERNARIO 5a Generación.v4Juan Carlos Diaz MartinezAún no hay calificaciones
- 8385-Texto Del Artículo-23420-1-10-20190906Documento14 páginas8385-Texto Del Artículo-23420-1-10-20190906Antonella MüllerAún no hay calificaciones
- El Almohadón de Plumas TrabajoDocumento7 páginasEl Almohadón de Plumas TrabajoBarrios Ana MaríaAún no hay calificaciones
- Excitable Cells: Monographs in Modern Biology for Upper School and University CoursesDe EverandExcitable Cells: Monographs in Modern Biology for Upper School and University CoursesAún no hay calificaciones
- Remapping Your Mind: The Neuroscience of Self-Transformation through StoryDe EverandRemapping Your Mind: The Neuroscience of Self-Transformation through StoryCalificación: 5 de 5 estrellas5/5 (2)
- The Synapse: Structure and FunctionDe EverandThe Synapse: Structure and FunctionVirginia M. PickelAún no hay calificaciones
- The Power of NeuroplasticityDe EverandThe Power of NeuroplasticityCalificación: 4.5 de 5 estrellas4.5/5 (32)
- Nerve Membranes: A Study of the Biological and Chemical Aspects of Neuron–Glia RelationshipsDe EverandNerve Membranes: A Study of the Biological and Chemical Aspects of Neuron–Glia RelationshipsAún no hay calificaciones
- The Neuronal Codes of the CerebellumDe EverandThe Neuronal Codes of the CerebellumDetlef HeckAún no hay calificaciones
- Create a Better Brain through Neuroplasticity: A Manual for MamasDe EverandCreate a Better Brain through Neuroplasticity: A Manual for MamasCalificación: 5 de 5 estrellas5/5 (1)
- The Biology of Thought: A Neuronal Mechanism in the Generation of Thought - A New Molecular ModelDe EverandThe Biology of Thought: A Neuronal Mechanism in the Generation of Thought - A New Molecular ModelCalificación: 5 de 5 estrellas5/5 (4)
- Nervous System: The Dog Stole the Professor's NotesDe EverandNervous System: The Dog Stole the Professor's NotesAún no hay calificaciones
- Behavioral Neuroscience: An IntroductionDe EverandBehavioral Neuroscience: An IntroductionCalificación: 5 de 5 estrellas5/5 (1)
- ¿Neurona Mata Espíritu? Una Aproximación Interdisciplinar al Cerebro, la Conciencia y la LibertadDe Everand¿Neurona Mata Espíritu? Una Aproximación Interdisciplinar al Cerebro, la Conciencia y la LibertadCalificación: 5 de 5 estrellas5/5 (1)
- Brain Training: Fascinating Tips For Memory Improvement, Concentration, Mental Clarity, Neuroplasticity, And Mind PowerDe EverandBrain Training: Fascinating Tips For Memory Improvement, Concentration, Mental Clarity, Neuroplasticity, And Mind PowerAún no hay calificaciones
- Developmental Plasticity: Behavioral and Biological Aspects of Variations in DevelopmentDe EverandDevelopmental Plasticity: Behavioral and Biological Aspects of Variations in DevelopmentEugene GollinAún no hay calificaciones
- Brain Training: The Ultimate Guide to Increase Your Brain Power and Improving Your Memory (Brain Exercise, Concentration, Neuroplasticity, Mental Clarity, Brain Plasticity)De EverandBrain Training: The Ultimate Guide to Increase Your Brain Power and Improving Your Memory (Brain Exercise, Concentration, Neuroplasticity, Mental Clarity, Brain Plasticity)Aún no hay calificaciones
- The Boy Who Could Run But Not Walk: Understanding Neuroplasticity in the Child's BrainDe EverandThe Boy Who Could Run But Not Walk: Understanding Neuroplasticity in the Child's BrainAún no hay calificaciones
- The Release of Catecholamines from Adrenergic NeuronsDe EverandThe Release of Catecholamines from Adrenergic NeuronsAún no hay calificaciones
- Neuronal and Synaptic Dysfunction in Autism Spectrum Disorder and Intellectual DisabilityDe EverandNeuronal and Synaptic Dysfunction in Autism Spectrum Disorder and Intellectual DisabilityCarlo SalaAún no hay calificaciones
- Acetylcholine: An Approach to the Molecular Mechanism of ActionDe EverandAcetylcholine: An Approach to the Molecular Mechanism of ActionAún no hay calificaciones
- How To Become Smarter: 21 Ways to Increase Your Brain Power in the Next 30 DaysDe EverandHow To Become Smarter: 21 Ways to Increase Your Brain Power in the Next 30 DaysAún no hay calificaciones
- Intracellular Consequences of Amyloid in Alzheimer's DiseaseDe EverandIntracellular Consequences of Amyloid in Alzheimer's DiseaseAún no hay calificaciones
- Brain Energetics and Neuronal Activity: Applications to fMRI and MedicineDe EverandBrain Energetics and Neuronal Activity: Applications to fMRI and MedicineAún no hay calificaciones
- Cellular Migration and Formation of Neuronal Connections: Comprehensive Developmental NeuroscienceDe EverandCellular Migration and Formation of Neuronal Connections: Comprehensive Developmental NeuroscienceCalificación: 5 de 5 estrellas5/5 (1)