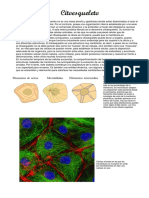
También podría gustarte
- Motilidad CelularDocumento4 páginasMotilidad CelularFrancis NicoleAún no hay calificaciones
- Orgánulos No Membranosos 2018Documento5 páginasOrgánulos No Membranosos 2018María Dolores Muñoz CastroAún no hay calificaciones
- Orgánulos No MembranososDocumento5 páginasOrgánulos No MembranososMaría Dolores Muñoz CastroAún no hay calificaciones
- Cito EsqueletoDocumento4 páginasCito EsqueletoMateo DuraznoAún no hay calificaciones
- Citosol, Comunicaciones y CitoesqueletoDocumento4 páginasCitosol, Comunicaciones y CitoesqueletoPaulaAún no hay calificaciones
- Estructuras y Organulos No MebranososDocumento7 páginasEstructuras y Organulos No MebranososRaul Ibanez RodriguezAún no hay calificaciones
- B2 T4 - CÉLULA EUCARIOTA - Citoplasma ApuntesDocumento16 páginasB2 T4 - CÉLULA EUCARIOTA - Citoplasma ApuntesNacho Orio FernándezAún no hay calificaciones
- Biología P19Documento15 páginasBiología P19jg2488955Aún no hay calificaciones
- Citoesqueleto transporte microtúbulosDocumento22 páginasCitoesqueleto transporte microtúbulosAndrésZonanaAún no hay calificaciones
- BQF - 1A, B - BIOLOGIA - U IIIbDocumento31 páginasBQF - 1A, B - BIOLOGIA - U IIIbNahomy BustillosAún no hay calificaciones
- Citoesqueleto y ciclo celularDocumento15 páginasCitoesqueleto y ciclo celularDanna Valentina MartinezAún no hay calificaciones
- Citoplasma y Substancias Ergáticas PDFDocumento25 páginasCitoplasma y Substancias Ergáticas PDFCristian Rene Ortiz CacheAún no hay calificaciones
- ENSAYO Citoesqueleto Karisbel SDocumento4 páginasENSAYO Citoesqueleto Karisbel Skarisbel suarezAún no hay calificaciones
- Apunte CitoesqueletoDocumento4 páginasApunte CitoesqueletoAliciaAún no hay calificaciones
- EL CITOesqueletoDocumento4 páginasEL CITOesqueletoDiana Del Pilar Chaustre DuranAún no hay calificaciones
- Citoesqueleto: Estructura y funciones clave de los microfilamentos, filamentos intermedios y microtúbulosDocumento5 páginasCitoesqueleto: Estructura y funciones clave de los microfilamentos, filamentos intermedios y microtúbulosErika Maribel BadilloAún no hay calificaciones
- MICROTUBULOSDocumento4 páginasMICROTUBULOSdaniel feijooAún no hay calificaciones
- Tema 8. Orgánulos CelularesDocumento16 páginasTema 8. Orgánulos CelularesMarco MatellánAún no hay calificaciones
- Citosol y CitoesqueletoDocumento5 páginasCitosol y CitoesqueletoDelia GrauAún no hay calificaciones
- BIOLOGÍA Tema 8 (Bloque 2)Documento5 páginasBIOLOGÍA Tema 8 (Bloque 2)Carolina De la peña fernandezAún no hay calificaciones
- Citosol y CitoesqueletoDocumento3 páginasCitosol y CitoesqueletosesechaAún no hay calificaciones
- Que Son Los Microtubulos Filamentos IntermediosDocumento6 páginasQue Son Los Microtubulos Filamentos IntermediosMarvinRiveraAún no hay calificaciones
- 5 - Estructuras Celulares No MembranosasDocumento3 páginas5 - Estructuras Celulares No MembranosasErika PerezAún no hay calificaciones
- CITOESQUELETODocumento20 páginasCITOESQUELETOElizabeth Garcia GuzmanAún no hay calificaciones
- 7 Biocelmol Citoesqueleto 2019Documento51 páginas7 Biocelmol Citoesqueleto 2019Gary MTAún no hay calificaciones
- Filamentos intermedios: estructura y funciónDocumento7 páginasFilamentos intermedios: estructura y funciónCesar A ArauzAún no hay calificaciones
- Organelos CelularesDocumento12 páginasOrganelos CelularesJulio Rz100% (3)
- Citoesqueleto: Microfilamentos Filamentos Intermedios MicrotúbulosDocumento10 páginasCitoesqueleto: Microfilamentos Filamentos Intermedios MicrotúbulosDian GAún no hay calificaciones
- Biología Tema 7 Célula Eucariota Estructuras y Orgánulos No MembranososDocumento4 páginasBiología Tema 7 Célula Eucariota Estructuras y Orgánulos No MembranososAnaAún no hay calificaciones
- CitoesqueletoDocumento21 páginasCitoesqueletoSoraida Sanchez ParraAún no hay calificaciones
- Citoesqueleto OriginalDocumento16 páginasCitoesqueleto OriginalfranciscoAún no hay calificaciones
- Unidad 07 Citosol y CitoesqueletoDocumento10 páginasUnidad 07 Citosol y CitoesqueletojvlpAún no hay calificaciones
- Taller 7 Biologia CelularDocumento4 páginasTaller 7 Biologia CelularLAURA SOFIA CACERES AGUASAún no hay calificaciones
- Citoesqueleto y MovimientoDocumento14 páginasCitoesqueleto y Movimientoandreaave100% (2)
- Apuntes Cit 3okDocumento20 páginasApuntes Cit 3okLesly Yomara Zuñiga EscobarAún no hay calificaciones
- AFPD6. Citoesqueleto 2020 I. Preguntas GuíaDocumento8 páginasAFPD6. Citoesqueleto 2020 I. Preguntas GuíaSelim Carlos Seguin AlfaroAún no hay calificaciones
- CITOESQUELETODocumento23 páginasCITOESQUELETOMeritxell Planell CastellsAún no hay calificaciones
- Unidad 9 CitoesqueletoDocumento43 páginasUnidad 9 CitoesqueletoAlba Fernández RevueltaAún no hay calificaciones
- Citoesqueleto TrabajoDocumento7 páginasCitoesqueleto TrabajoNenita HermoxaAún no hay calificaciones
- Célula EucariotaDocumento5 páginasCélula EucariotaSara VeraAún no hay calificaciones
- Biologia MitocondriasDocumento7 páginasBiologia MitocondriasMELANI BRIGHIT RIVAS BUSTEAún no hay calificaciones
- Membranas y estructuras de los cloroplastosDocumento8 páginasMembranas y estructuras de los cloroplastosEstebin PaterninaAún no hay calificaciones
- Citoplasma y citoesqueleto: componentes y funcionesDocumento50 páginasCitoplasma y citoesqueleto: componentes y funcionesyaritzaAún no hay calificaciones
- Seg Exa BiologiaDocumento36 páginasSeg Exa BiologiaBrayan Villavicencio TovarAún no hay calificaciones
- Que Es El ProtoplasmaDocumento13 páginasQue Es El ProtoplasmaDenian CastilloAún no hay calificaciones
- El citoesqueleto: microfilamentos, microtúbulos y filamentos intermediosDocumento5 páginasEl citoesqueleto: microfilamentos, microtúbulos y filamentos intermediossandraAún no hay calificaciones
- wuolah-free-app-1707050491591Documento8 páginaswuolah-free-app-1707050491591marinagarciapalomaresAún no hay calificaciones
- HELENA CURTIS. Capítulo 2 de CÉLULASDocumento3 páginasHELENA CURTIS. Capítulo 2 de CÉLULASVane DiazAún no hay calificaciones
- Citoesqueleto CelularDocumento2 páginasCitoesqueleto Celular18388919Aún no hay calificaciones
- Estructura y funciones del citoesqueletoDocumento32 páginasEstructura y funciones del citoesqueletoGabriela IbañezAún no hay calificaciones
- Citoesqueleto Final IntroDocumento6 páginasCitoesqueleto Final IntroBelenMinAún no hay calificaciones
- CITOESKELETODocumento14 páginasCITOESKELETOJose Vicente Banda NuñezAún no hay calificaciones
- Unidad Siete CitoesqueletoDocumento57 páginasUnidad Siete CitoesqueletoJoe Mario ChavezAún no hay calificaciones
- Tema 08 Citosol y orgánulos no membranosos de la célulaDocumento16 páginasTema 08 Citosol y orgánulos no membranosos de la célulacandelaropero2006Aún no hay calificaciones
- Organelos celulares: estructuras y funcionesDocumento74 páginasOrganelos celulares: estructuras y funcionesRaikmg martinAún no hay calificaciones
- Caracter Í SticasDocumento19 páginasCaracter Í Sticashanna.aldanaAún no hay calificaciones
- Núcleo y CitoesqueletoDocumento2 páginasNúcleo y CitoesqueletoMelisa Mtz. Quijano.Aún no hay calificaciones
- Copia de Documento de PauDocumento11 páginasCopia de Documento de Paupmvm2004Aún no hay calificaciones
- Neurobiologia de PsicoterapiasDocumento19 páginasNeurobiologia de PsicoterapiasmarigeltbAún no hay calificaciones
- Aparato reproductor masculino: anatomía y fisiologíaDocumento143 páginasAparato reproductor masculino: anatomía y fisiologíaXimena Cisneros A.Aún no hay calificaciones
- 4 - UrinarioDocumento17 páginas4 - UrinarioXilena SilvaAún no hay calificaciones
- Anatomía y fisiología sistemas reproductoresDocumento26 páginasAnatomía y fisiología sistemas reproductoresDavid Alvarez VillotaAún no hay calificaciones
- Hemerobius BolivariDocumento4 páginasHemerobius BolivariRonnald HuallpaAún no hay calificaciones
- Mapas FiDocumento3 páginasMapas Fimaleja HernándezAún no hay calificaciones
- Prueba de La Unidad 3. Sistema Nervioso y Endocrino UNALMDocumento7 páginasPrueba de La Unidad 3. Sistema Nervioso y Endocrino UNALMRusvel Alex Vilca SivirichiAún no hay calificaciones
- Hormonas Del Hipotálamo y de La HipófisisDocumento8 páginasHormonas Del Hipotálamo y de La HipófisisLESTER OSIEL PEREZ OROZCOAún no hay calificaciones
- Estructuras y procesos de reproducción en hongosDocumento12 páginasEstructuras y procesos de reproducción en hongosximena MalrauxAún no hay calificaciones
- Ficha Tecnica TROLVIT AMINOACIDOSDocumento4 páginasFicha Tecnica TROLVIT AMINOACIDOSMilagros Baquerizo HuamánAún no hay calificaciones
- Tema 13. Orden KinetoplastidaDocumento11 páginasTema 13. Orden KinetoplastidaarianavaleroulaAún no hay calificaciones
- Tratamiento Específico para EndometriosisDocumento3 páginasTratamiento Específico para EndometriosisMariela AlvarezAún no hay calificaciones
- La Neurona: Potencial de Membrana y Potencial de AcciónDocumento14 páginasLa Neurona: Potencial de Membrana y Potencial de AcciónReycel Lapierre PobleteAún no hay calificaciones
- Voz NasalDocumento1 páginaVoz NasalIgnacio Nicolas Diaz SotoAún no hay calificaciones
- EMBRIOLOGIADocumento17 páginasEMBRIOLOGIASary MTAún no hay calificaciones
- El Sistema Nervioso Es Uno de Los Más Importantes y Complejos Del Cuerpo HumanoDocumento13 páginasEl Sistema Nervioso Es Uno de Los Más Importantes y Complejos Del Cuerpo HumanoOsiris Daniela Cisnados AvilaAún no hay calificaciones
- La influencia de la felicidad en el sistema cardiovascularDocumento17 páginasLa influencia de la felicidad en el sistema cardiovascularSonia AsensioAún no hay calificaciones
- Patokinesis ArtrosisDocumento1 páginaPatokinesis ArtrosisValentina G. GamboaAún no hay calificaciones
- Relación farmacología cienciasDocumento1 páginaRelación farmacología cienciasScarlet Geraldo100% (1)
- Transporte A Traves de MembranaDocumento25 páginasTransporte A Traves de MembranaSharon Cubas QuezadaAún no hay calificaciones
- Datos Acerca de La PielDocumento40 páginasDatos Acerca de La PielDulce SoledadAún no hay calificaciones
- 53b330dd30329 - Ficha Tecnica CitoneuronDocumento4 páginas53b330dd30329 - Ficha Tecnica Citoneuronjennifer racagni espinozaAún no hay calificaciones
- 07 Anabolizantes Beneficios e MaleficiosDocumento2 páginas07 Anabolizantes Beneficios e MaleficiosKlysmann ReisAún no hay calificaciones
- Pares Alineacion ColumnaDocumento4 páginasPares Alineacion Columnamisabelfuente3164Aún no hay calificaciones
- AGA ULTIMO Actual - PptxhoyyyyDocumento27 páginasAGA ULTIMO Actual - PptxhoyyyyLuis Perez RiquelmeAún no hay calificaciones
- Psicoprofilaxis de EmergenciaDocumento10 páginasPsicoprofilaxis de Emergencia05-OB-HU-ADALIZ SILVIA CERRON LAZOAún no hay calificaciones
- TAREA1-B1 Mapa Mental Del Sistema Nervioso Central y Sus Funciones - RemovedDocumento2 páginasTAREA1-B1 Mapa Mental Del Sistema Nervioso Central y Sus Funciones - RemovedAngelica RodríguezAún no hay calificaciones
- Bases Biologicas de La ConductaDocumento7 páginasBases Biologicas de La ConductaFabry EspinalAún no hay calificaciones
- Naturales de QuintoDocumento3 páginasNaturales de QuintoCristian D. Reyes VillarrealAún no hay calificaciones
- CMR Español 2 PDFDocumento14 páginasCMR Español 2 PDFMauricio LedesmaAún no hay calificaciones