También podría gustarte
- Mejoramiento genético de plantas: Segunda EdiciónDe EverandMejoramiento genético de plantas: Segunda EdiciónCalificación: 3 de 5 estrellas3/5 (5)
- Manual De Buenas Prácticas Ganaderas En Unidades De Producción Que Participan En El Corredor Pacífico Sur Guerrerense.De EverandManual De Buenas Prácticas Ganaderas En Unidades De Producción Que Participan En El Corredor Pacífico Sur Guerrerense.Aún no hay calificaciones
- Genética cuantitativa aplicada al mejoramiento animalDe EverandGenética cuantitativa aplicada al mejoramiento animalCalificación: 4 de 5 estrellas4/5 (4)
- La menarquia y las fases del ciclo menstrualDocumento3 páginasLa menarquia y las fases del ciclo menstrualDante Mendoza Lama90% (10)
- El búfalo de agua. Tomo 2: Generalidades y características productivasDe EverandEl búfalo de agua. Tomo 2: Generalidades y características productivasAún no hay calificaciones
- Nuevos horizontes del mejoramiento genético animalDocumento25 páginasNuevos horizontes del mejoramiento genético animalYesid L. Huanca Quito100% (1)
- Transferencia de Embriones en Ganado BovinoDocumento16 páginasTransferencia de Embriones en Ganado BovinoVenezuelaGanadera.comAún no hay calificaciones
- Introduccion Al Mejoramiento Genetico AnimalDocumento74 páginasIntroduccion Al Mejoramiento Genetico AnimalOliver Hato100% (2)
- Seleccion de PorcinosDocumento5 páginasSeleccion de PorcinosFrancisco SalgueroAún no hay calificaciones
- Resumen de Mejoramiento GeneticoDocumento12 páginasResumen de Mejoramiento GeneticoJhino Orlando Ventura AlvarezAún no hay calificaciones
- 1 Era Clase GeneticaDocumento24 páginas1 Era Clase GeneticaCarlos MartinezAún no hay calificaciones
- Herramienta para Un Crecimiento Permanente: Mejoramiento Genético AnimalDocumento4 páginasHerramienta para Un Crecimiento Permanente: Mejoramiento Genético AnimalSebastian Martinez RodriguezAún no hay calificaciones
- Ovinos Paramétros GenéticosDocumento28 páginasOvinos Paramétros Genéticosijbinijchipichipi20Aún no hay calificaciones
- Genetica Porcigenes-64196f200972dDocumento11 páginasGenetica Porcigenes-64196f200972dHelen QuinteroAún no hay calificaciones
- Mejoramiento Genetico - UNALMDocumento36 páginasMejoramiento Genetico - UNALMAngel Vasqz RAún no hay calificaciones
- Articulos Unidad 4Documento24 páginasArticulos Unidad 4AlbaYanethCortesEstevezAún no hay calificaciones
- GENOMICADocumento4 páginasGENOMICAMarysabel Vicuña CardenasAún no hay calificaciones
- Métodos de mejora genética sostenible en recursos zoogenéticosDocumento27 páginasMétodos de mejora genética sostenible en recursos zoogenéticosRaul LopezAún no hay calificaciones
- Aspectos Generales de Mejoramiento GenéticoDocumento14 páginasAspectos Generales de Mejoramiento GenéticoDavid Guillermo Merchancano BravoAún no hay calificaciones
- Calculo Heredabilidad y Varianza en Un Plantel Porcino. Yaz 1Documento28 páginasCalculo Heredabilidad y Varianza en Un Plantel Porcino. Yaz 1Joffre SaulAún no hay calificaciones
- Jeb207159 FuDocumento18 páginasJeb207159 FuCamilo DAún no hay calificaciones
- Componentes del mejoramiento genético en ovino y caprinoDocumento9 páginasComponentes del mejoramiento genético en ovino y caprinoaura aularAún no hay calificaciones
- 97-Texto Del Artículo-266-1-10-20131222Documento7 páginas97-Texto Del Artículo-266-1-10-20131222Sergio MolinaAún no hay calificaciones
- Mejoramiento genético de cuyes para incrementar la producciónDocumento19 páginasMejoramiento genético de cuyes para incrementar la producciónWilliam V. Anahua100% (1)
- Avances Del Mejoramiento Genetico en ConejosDocumento8 páginasAvances Del Mejoramiento Genetico en ConejosIswelly AlvaradoAún no hay calificaciones
- Metodos de Mejora GeneticaDocumento52 páginasMetodos de Mejora GeneticaAntonio Fernandez JimenezAún no hay calificaciones
- Curso de Mejoramiento GeneticoDocumento45 páginasCurso de Mejoramiento GeneticoJhonny Roque HuaracaAún no hay calificaciones
- Biotecnología Aplicada A La Ganadería MonografiaDocumento21 páginasBiotecnología Aplicada A La Ganadería MonografiaSoledad VegaAún no hay calificaciones
- Heredabilidad - Del CerdoDocumento5 páginasHeredabilidad - Del CerdoMagui CendejasAún no hay calificaciones
- Biotecnología y Mejoramiento Genético en Ovinos y CaprinosDocumento3 páginasBiotecnología y Mejoramiento Genético en Ovinos y CaprinosBrandy BenjaminAún no hay calificaciones
- Mejoramiento Genetico BovinoDocumento36 páginasMejoramiento Genetico Bovinoemp.com.ecoljaAún no hay calificaciones
- Genética Molecular para El Mejoramiento de Los Rasgos Reproductivos en Bovinos GENETICADocumento9 páginasGenética Molecular para El Mejoramiento de Los Rasgos Reproductivos en Bovinos GENETICAVenezuelaGanadera.comAún no hay calificaciones
- NR38438Documento3 páginasNR38438Ronald Rosado SaltosAún no hay calificaciones
- Estimación de Heterosis en CuyesDocumento13 páginasEstimación de Heterosis en CuyesElmer Meza RAún no hay calificaciones
- 1Documento7 páginas1Edison torres alvarezAún no hay calificaciones
- Universidad Nacional Del Altiplano Puno GeneticaDocumento10 páginasUniversidad Nacional Del Altiplano Puno GeneticaNelson CariAún no hay calificaciones
- Mej. Animal 1Documento77 páginasMej. Animal 1hectorAún no hay calificaciones
- NR30037Documento9 páginasNR30037Carlos Alberto Artica PinedaAún no hay calificaciones
- 2 Mejoramiento Genético de Aves y CerdosDocumento3 páginas2 Mejoramiento Genético de Aves y CerdosNoemí Paz Matamala GarcíaAún no hay calificaciones
- SELECCIONDocumento24 páginasSELECCIONLuis Jose Rivero RiveroAún no hay calificaciones
- 2010 Mejora Abejas 2 El Colmenar PDFDocumento8 páginas2010 Mejora Abejas 2 El Colmenar PDFNilo C Cervantes ChipaAún no hay calificaciones
- Manual Mejoramiento GenéticoDocumento86 páginasManual Mejoramiento GenéticoSAMARA GONZALEZ NU�EZAún no hay calificaciones
- Dialnet AspectosDeLaGeneticaMolecularCunicola1Parte 2869767Documento9 páginasDialnet AspectosDeLaGeneticaMolecularCunicola1Parte 2869767Favio AmaqAún no hay calificaciones
- 2.1.2 Eficiencia ReproductivaDocumento16 páginas2.1.2 Eficiencia ReproductivaDiana ValenzuelaAún no hay calificaciones
- Biologia Reproduccion PalmaDocumento35 páginasBiologia Reproduccion PalmaHenry MVZAún no hay calificaciones
- Fundamentos básicos de genética cuantitativa en el contexto del mejoramiento genético animalDocumento44 páginasFundamentos básicos de genética cuantitativa en el contexto del mejoramiento genético animalElmer Meza RojasAún no hay calificaciones
- Mejoramiento GeneticoDocumento10 páginasMejoramiento GeneticoNicasio RodríguezAún no hay calificaciones
- Propuesta Yadilxa (Autoguardado)Documento17 páginasPropuesta Yadilxa (Autoguardado)José NuñezAún no hay calificaciones
- PigCHAMP Articulos - Granjas de Genética: Particularidades de Manejo y Gestión de DatosDocumento5 páginasPigCHAMP Articulos - Granjas de Genética: Particularidades de Manejo y Gestión de DatosPigCHAMP_Pro_EuropaAún no hay calificaciones
- CLASE 1 REPRODUCCIÒN ANIMAL Marzo2020 PDFDocumento31 páginasCLASE 1 REPRODUCCIÒN ANIMAL Marzo2020 PDFCristhian MoralesAún no hay calificaciones
- Genética Clase 1Documento3 páginasGenética Clase 1Liz suntaxiAún no hay calificaciones
- Estrategias de Mejoramiento GeneticoDocumento11 páginasEstrategias de Mejoramiento GeneticoWilberto Cruz PastoraAún no hay calificaciones
- A17704 PDFDocumento4 páginasA17704 PDFRuben SerraAún no hay calificaciones
- 5 CC 204 BBD 8087Documento14 páginas5 CC 204 BBD 8087Marianyelis bermudezAún no hay calificaciones
- NR38526 PDFDocumento15 páginasNR38526 PDFefrainAún no hay calificaciones
- Catalogo Bovinos Cebu Agosto2018Documento82 páginasCatalogo Bovinos Cebu Agosto2018Boettiger Zepeda Emilia 5Aún no hay calificaciones
- Ganado lechero: Importancia de la interacción genotipo-ambienteDocumento7 páginasGanado lechero: Importancia de la interacción genotipo-ambienteAletheia1708Aún no hay calificaciones
- 26 29 Limites Seleccion Genetica Avicultura Teoricos Biologicos Eticos Ambiental SA201810Documento4 páginas26 29 Limites Seleccion Genetica Avicultura Teoricos Biologicos Eticos Ambiental SA201810Karen Contreras JaimesAún no hay calificaciones
- Mejoramiento Genetico en AcuiculturaDocumento8 páginasMejoramiento Genetico en AcuiculturaAlejandra ContrerasAún no hay calificaciones
- Aplicaciones de biotecnología y bioinformática en diferentes áreas de la producción animalDe EverandAplicaciones de biotecnología y bioinformática en diferentes áreas de la producción animalAún no hay calificaciones
- Etapas Del EmbarazoDocumento9 páginasEtapas Del EmbarazoFavian AlmanzaAún no hay calificaciones
- Instructivo para el tamizaje de violencia en niñas y adolescentesDocumento51 páginasInstructivo para el tamizaje de violencia en niñas y adolescentesEuler LupercioAún no hay calificaciones
- Clases de La Semana Del 11-05-2020 Al 15-05-2020 Docente Vinayde Duque Proyecto: Familia y Colegio Unidos Vía Digital Por Mi Educación MatemáticaDocumento3 páginasClases de La Semana Del 11-05-2020 Al 15-05-2020 Docente Vinayde Duque Proyecto: Familia y Colegio Unidos Vía Digital Por Mi Educación MatemáticaJahannay Sley Fajardo JaimesAún no hay calificaciones
- Doble Penetración - Wikipedia, La Enciclopedia Libre PDFDocumento18 páginasDoble Penetración - Wikipedia, La Enciclopedia Libre PDFSam NarváezAún no hay calificaciones
- Ii BimestreDocumento35 páginasIi BimestreJoscimar Gamarra MejíaAún no hay calificaciones
- Exploracion Fisica Del Aparato Femenino Miranda Becerril Edgar 5HM5Documento22 páginasExploracion Fisica Del Aparato Femenino Miranda Becerril Edgar 5HM5Miranda Becerril Edgar JonathanAún no hay calificaciones
- DebateDocumento2 páginasDebateFelipe HidalgoAún no hay calificaciones
- Reproduccion AnimalDocumento32 páginasReproduccion AnimalMaría Gabriela ArangurenAún no hay calificaciones
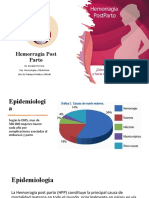
- Hemorragia Post PartoDocumento34 páginasHemorragia Post PartogastonAún no hay calificaciones
- Consejería en planificación familiar en adolescentesDocumento79 páginasConsejería en planificación familiar en adolescentesLeticia ConstantinoAún no hay calificaciones
- Fallo Osde Reproducción AsistidaDocumento17 páginasFallo Osde Reproducción AsistidaEntre Ríos AhoraAún no hay calificaciones
- CasoObstetriciaDolorEpigastrioDocumento9 páginasCasoObstetriciaDolorEpigastrioCamila AcevedoAún no hay calificaciones
- Contactos Mujeres Griego Barato en MadridDocumento2 páginasContactos Mujeres Griego Barato en MadridEncontrar pareja en MadridAún no hay calificaciones
- Hipogonadismo - Incluye HH, KlinefelterDocumento20 páginasHipogonadismo - Incluye HH, KlinefelterGabrielaAún no hay calificaciones
- Hemorragias de La Primera Mitad Del EmbarazoDocumento12 páginasHemorragias de La Primera Mitad Del EmbarazoAlmaAún no hay calificaciones
- 4to Grado - Bloque 1 - Ejercicios ComplementariosDocumento26 páginas4to Grado - Bloque 1 - Ejercicios Complementariosmiriam230989% (9)
- Resumen Cambios Fisiológicos y Anatómicos Durante La GestaciónDocumento6 páginasResumen Cambios Fisiológicos y Anatómicos Durante La GestaciónSamuel Espitia CastroAún no hay calificaciones
- CARTA DESCRIPTIVA PaapanicolauDocumento7 páginasCARTA DESCRIPTIVA PaapanicolauJavier G Torrico M100% (1)
- Anatomía del aparato reproductor macho y hembraDocumento33 páginasAnatomía del aparato reproductor macho y hembraYURANI VAZQUEZ AGUILERAAún no hay calificaciones
- Proliferación ovogonias ovarioDocumento8 páginasProliferación ovogonias ovarioyolimar loretoAún no hay calificaciones
- Informe de Sistematizacion Sexualidad Adolescente Desde El TsDocumento16 páginasInforme de Sistematizacion Sexualidad Adolescente Desde El TsMicaela MuñozAún no hay calificaciones
- Aparatos del cuerpo humano y sus funcionesDocumento5 páginasAparatos del cuerpo humano y sus funcionesDarwin BeckhamAún no hay calificaciones
- Efectos de LevonorgestrelDocumento2 páginasEfectos de LevonorgestrelCinjul AriasAún no hay calificaciones
- Hilchos Mechussarey KapparahDocumento14 páginasHilchos Mechussarey KapparahIojanan Ovadiah CastroAún no hay calificaciones
- Mais 2018Documento214 páginasMais 2018perezitalo593Aún no hay calificaciones
- Disfunción Ovárica (Mariersy)Documento8 páginasDisfunción Ovárica (Mariersy)GERAL FERNANDEZAún no hay calificaciones
- Protocolo Diagnostico en Amenorrea PrimariaDocumento10 páginasProtocolo Diagnostico en Amenorrea PrimariaJose Alejandro Silva SaavedraAún no hay calificaciones
- Protocolo de Órdenes de Ginecologia y ObstetriciaDocumento10 páginasProtocolo de Órdenes de Ginecologia y ObstetriciaJohana Contreras100% (2)
- Mehu131 U2 T7 ReproduccionfemeninaDocumento30 páginasMehu131 U2 T7 ReproduccionfemeninaSheila EsRaAún no hay calificaciones