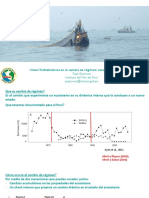
También podría gustarte
- Dupuy Et Al. 1993. Estructura Del Bosque Enano Nublado Macuira PDFDocumento6 páginasDupuy Et Al. 1993. Estructura Del Bosque Enano Nublado Macuira PDFDaniela PiraquiveAún no hay calificaciones
- Flora y Vegetacion de La Provincia Arque, Departamento Cochabamba, BoliviaDocumento14 páginasFlora y Vegetacion de La Provincia Arque, Departamento Cochabamba, BoliviaCristian EverthAún no hay calificaciones
- Diversidad taxonómica y funcional de lianas en bosque tropical del ChocóDocumento29 páginasDiversidad taxonómica y funcional de lianas en bosque tropical del ChocóJohana CopeteAún no hay calificaciones
- Tema 26. Evol HominidosDocumento88 páginasTema 26. Evol HominidosJuanma Fernandez RatiaAún no hay calificaciones
- Fotosintesisproductividadyalgasmarinas Eddingetal2006Documento40 páginasFotosintesisproductividadyalgasmarinas Eddingetal2006EgofrutAún no hay calificaciones
- Waisfeld Vaccari 2008 BioestratiDocumento10 páginasWaisfeld Vaccari 2008 BioestratiwillymantorAún no hay calificaciones
- Analisis Multivariado AplicadoDocumento350 páginasAnalisis Multivariado AplicadoAnonymous MVQLe29anXAún no hay calificaciones
- CofechadoDocumento32 páginasCofechadoLuis David HuaytaAún no hay calificaciones
- Morfometría, Abundancia y Distribución de Ucides Occidentalis en El Sector Occidental de La Bahía de JiquiliscoDocumento19 páginasMorfometría, Abundancia y Distribución de Ucides Occidentalis en El Sector Occidental de La Bahía de JiquiliscoCarlos RiveraAún no hay calificaciones
- Climent-Domènech.H - 2008 - An Example of Paleoecological Analysis Using The Cluster Method. Bartonian Reef Facies (Vic, NE From The Ebro Basin)Documento4 páginasCliment-Domènech.H - 2008 - An Example of Paleoecological Analysis Using The Cluster Method. Bartonian Reef Facies (Vic, NE From The Ebro Basin)Hasdrúbal Climent-DomènechAún no hay calificaciones
- Lopez-Rasgado - Semana Posgrado2012Documento17 páginasLopez-Rasgado - Semana Posgrado2012FRANCISCO RASGADOAún no hay calificaciones
- Tecnicas Fitoplancton Jorge Bixler PDFDocumento27 páginasTecnicas Fitoplancton Jorge Bixler PDFGeeorge BixleerAún no hay calificaciones
- Proyecto Hidras, Final ZoologiaDocumento15 páginasProyecto Hidras, Final ZoologiaJuanjo VillalobosAún no hay calificaciones
- Tecnicas Fitoplancton Jorge BixlerDocumento27 páginasTecnicas Fitoplancton Jorge BixlerGeeorge BixleerAún no hay calificaciones
- Rehabilitación del recinto del tapirDocumento32 páginasRehabilitación del recinto del tapirRuth Cabello HerreraAún no hay calificaciones
- 54014203Documento7 páginas54014203Jonathan MarchánAún no hay calificaciones
- 405Documento819 páginas405Stefania MeraAún no hay calificaciones
- 3 Cve331 c3 PptvideoconferenciaDocumento44 páginas3 Cve331 c3 PptvideoconferenciaValentina JaraAún no hay calificaciones
- PPT-Rildo Rojas TuanamaDocumento28 páginasPPT-Rildo Rojas TuanamaMauricio Royer Mamani CruzAún no hay calificaciones
- Ecología AcuáticaDocumento15 páginasEcología Acuáticaxiadani gutierrezAún no hay calificaciones
- Ciclo AndinoDocumento64 páginasCiclo AndinoByron Omar Zambrano MedinaAún no hay calificaciones
- Rocky Reef Benthic Assemblages in The Magellan Strait and The South Shetland Islands (Antarctica) Newcombe & Cárdenas 2011Documento12 páginasRocky Reef Benthic Assemblages in The Magellan Strait and The South Shetland Islands (Antarctica) Newcombe & Cárdenas 2011Catalina Merino YunnissiAún no hay calificaciones
- Nelson ZamoraDocumento69 páginasNelson ZamoraEimyAún no hay calificaciones
- Cap 2 Ecorregiones PunaDocumento37 páginasCap 2 Ecorregiones PunaJohanna AyalaAún no hay calificaciones
- Conferencia Dr. Pepe Espinoza SilveraDocumento33 páginasConferencia Dr. Pepe Espinoza SilveraCesar LapaAún no hay calificaciones
- 0306 Divers i Dad de Organism OsDocumento110 páginas0306 Divers i Dad de Organism OsYue CaeruleaAún no hay calificaciones
- Presentacion ClasesDocumento42 páginasPresentacion ClasesCantoJAún no hay calificaciones
- Clase ComunidadDocumento34 páginasClase ComunidadNicolas WAún no hay calificaciones
- Practica 1Documento3 páginasPractica 1andersonalexanderespinozaAún no hay calificaciones
- Guia Macroalgas Costa Mich 2008-1Documento64 páginasGuia Macroalgas Costa Mich 2008-1larissaAún no hay calificaciones
- Identificacionde Corales TropicalesDocumento63 páginasIdentificacionde Corales TropicalesComo MuchoAún no hay calificaciones
- Articulo Pro-Explo 2015 Victor Valdivia PDFDocumento6 páginasArticulo Pro-Explo 2015 Victor Valdivia PDFVictor ValdiviaAún no hay calificaciones
- Actividad Corrientes OceanicasDocumento3 páginasActividad Corrientes OceanicasCatalina Contreras YanténAún no hay calificaciones
- Vasquez Vega BIODIVERSIDAD-U-CHDocumento28 páginasVasquez Vega BIODIVERSIDAD-U-CHKATALINA IGNACIA AZÓCARAún no hay calificaciones
- Check_rapposa_extensión Austral a La Distribución Conocida de MicoureusDocumento4 páginasCheck_rapposa_extensión Austral a La Distribución Conocida de Micoureusgbitencourt.bioAún no hay calificaciones
- 1 - Concepto, Clima, Hidrología y Eventos ExtremosDocumento32 páginas1 - Concepto, Clima, Hidrología y Eventos ExtremosFer Absolem ChipanaAún no hay calificaciones
- La ecología: la ciencia de las interaccionesDocumento46 páginasLa ecología: la ciencia de las interaccionesCatalina Merino YunnissiAún no hay calificaciones
- Sillar Geologia Regional 5°entreDocumento27 páginasSillar Geologia Regional 5°entrejuan antonioAún no hay calificaciones
- Aplicaciones de Las Datación Por Series de Uranio en Paleontología de InvertebradosDocumento11 páginasAplicaciones de Las Datación Por Series de Uranio en Paleontología de InvertebradosManuel Reyna PahuachoAún no hay calificaciones
- Salida campo Ancón recursos hidrobiológicosDocumento25 páginasSalida campo Ancón recursos hidrobiológicosdemiAún no hay calificaciones
- Abdala y Diaz 2000 Akodon AlbiventerDocumento18 páginasAbdala y Diaz 2000 Akodon Albiventerruthmcontreras88Aún no hay calificaciones
- ÌNDICE DE DiversidadDocumento15 páginasÌNDICE DE DiversidadKarina TamayoAún no hay calificaciones
- Instructivo 3. Comunidades.Documento33 páginasInstructivo 3. Comunidades.PABLO OSWALDO JHAIR PACHECO ROJASAún no hay calificaciones
- Diversidad Alfa y Beta en Bosque seco-ANMI MADIDI-BOLIVIADocumento19 páginasDiversidad Alfa y Beta en Bosque seco-ANMI MADIDI-BOLIVIAdianaAún no hay calificaciones
- Codigo Estratigrafico NorteamericanoDocumento8 páginasCodigo Estratigrafico NorteamericanoIxchel Yolotzin Apodaca MoraAún no hay calificaciones
- Variación Anatómica de La Madera de Acacia Melanoxylon Implantada en El Sudeste de La Provincia de Buenos AiresDocumento7 páginasVariación Anatómica de La Madera de Acacia Melanoxylon Implantada en El Sudeste de La Provincia de Buenos AiresIvan LopezAún no hay calificaciones
- Estimación del tamaño poblacional de insectosDocumento5 páginasEstimación del tamaño poblacional de insectosEDUARDO MARTIN LOPEZ CORDOVAAún no hay calificaciones
- Serpientes de La Región Biogeográfica Del ChacoDocumento38 páginasSerpientes de La Región Biogeográfica Del ChacoSuri CabreraAún no hay calificaciones
- Evapofacies Del Salar Hombre Muerto Puna ArgentinaDocumento13 páginasEvapofacies Del Salar Hombre Muerto Puna ArgentinaLula FreijoAún no hay calificaciones
- PRACTICO 4 - Elasticidad 2019Documento7 páginasPRACTICO 4 - Elasticidad 2019roAún no hay calificaciones
- Patrones de Distribución de Macro InvertebradosDocumento18 páginasPatrones de Distribución de Macro InvertebradosalexAún no hay calificaciones
- Importancia de La Icnología en La Determinación de Ambientes Sedimentarios y La EstratigrafíaDocumento7 páginasImportancia de La Icnología en La Determinación de Ambientes Sedimentarios y La EstratigrafíakototokoAún no hay calificaciones
- El papel de la matriz en la conservación de la biodiversidad en paisajes fragmentadosDocumento14 páginasEl papel de la matriz en la conservación de la biodiversidad en paisajes fragmentadosKevin Arana RabanalAún no hay calificaciones
- Curvas de AcumulaciónDocumento6 páginasCurvas de AcumulaciónsaaAún no hay calificaciones
- Gasteropodos Marinos ColumbretesDocumento39 páginasGasteropodos Marinos ColumbretesPerico PalotesAún no hay calificaciones
- InformeDocumento39 páginasInformePandero CuscoAún no hay calificaciones
- Clase 3. Fósiles Guía, Bioestratigrafía y Carta CronoestratifDocumento26 páginasClase 3. Fósiles Guía, Bioestratigrafía y Carta CronoestratifMaximiliano RicoAún no hay calificaciones
- Libélulas y caballitos del diablo del departamento del Meta, ColombiaDe EverandLibélulas y caballitos del diablo del departamento del Meta, ColombiaAún no hay calificaciones
- Atlas de ciliados y otros microorganismos frecuentes en plantas de sistemas aerobio de aguas residualesDe EverandAtlas de ciliados y otros microorganismos frecuentes en plantas de sistemas aerobio de aguas residualesAún no hay calificaciones
- Emergencia de extinción animal: una aventura de Max Axiom, súpercientíficoDe EverandEmergencia de extinción animal: una aventura de Max Axiom, súpercientíficoAún no hay calificaciones
- 11 EvaluaciónDocumento4 páginas11 EvaluaciónCatalina Merino YunnissiAún no hay calificaciones
- Clase8 - Parasitismo, Enfermedades y CompetenciaDocumento40 páginasClase8 - Parasitismo, Enfermedades y CompetenciaCatalina Merino YunnissiAún no hay calificaciones
- La ecología: la ciencia de las interaccionesDocumento46 páginasLa ecología: la ciencia de las interaccionesCatalina Merino YunnissiAún no hay calificaciones
- Insta Laci OnDocumento1 páginaInsta Laci OnCatalina Merino YunnissiAún no hay calificaciones
- Clase9a - DepredaciónDocumento14 páginasClase9a - DepredaciónCatalina Merino YunnissiAún no hay calificaciones
- Clase 7 Metapoblaciones: Conceptos clave y modelos básicosDocumento35 páginasClase 7 Metapoblaciones: Conceptos clave y modelos básicosCatalina Merino YunnissiAún no hay calificaciones
- Estructura de edades en poblacionesDocumento39 páginasEstructura de edades en poblacionesCatalina Merino YunnissiAún no hay calificaciones
- Seguridad vial: frenos y distanciasDocumento5 páginasSeguridad vial: frenos y distanciasCatalina Merino YunnissiAún no hay calificaciones
- 2019 Acosta & Pérez González - Quién Va A Describir Nuestra Biodiversidad... Rev. Mus. Argentino Cienc. Nat., N.S. 21 (1), 17-27Documento11 páginas2019 Acosta & Pérez González - Quién Va A Describir Nuestra Biodiversidad... Rev. Mus. Argentino Cienc. Nat., N.S. 21 (1), 17-27Abel Perez GonzalezAún no hay calificaciones
- 1083-Texto Del Artículo-2847-1-10-20180405Documento45 páginas1083-Texto Del Artículo-2847-1-10-20180405Belén ZambranoAún no hay calificaciones
- Las Colecciones Científicas y La Protección Del Patrimonio Paleontológico.Documento7 páginasLas Colecciones Científicas y La Protección Del Patrimonio Paleontológico.López LuisAún no hay calificaciones
- Resumen Estudio - Propuesta Sistema de Gobhernanza de Género - SERPATDocumento10 páginasResumen Estudio - Propuesta Sistema de Gobhernanza de Género - SERPATCatalina Merino YunnissiAún no hay calificaciones
- Clase 4Documento63 páginasClase 4Catalina Merino YunnissiAún no hay calificaciones
- Clase 2Documento70 páginasClase 2Catalina Merino YunnissiAún no hay calificaciones
- XXIX JAM Taller - Principios Curatoriales para Las Colecciones de Historia NaturalDocumento3 páginasXXIX JAM Taller - Principios Curatoriales para Las Colecciones de Historia NaturalCatalina Merino YunnissiAún no hay calificaciones
- Zoogeografia de Los Crustaceos Decapodos ChilenosDocumento28 páginasZoogeografia de Los Crustaceos Decapodos ChilenosCatalina Merino YunnissiAún no hay calificaciones
- Colecciones Biológicas Una Alternativa PDFDocumento13 páginasColecciones Biológicas Una Alternativa PDFmayra garzon vargasAún no hay calificaciones
- Agentes DeterioroDocumento47 páginasAgentes DeterioroCatalina Merino YunnissiAún no hay calificaciones
- SPATIAL DISTRIBUTION AND RELATIVE ABUNDANCE OF ECHINODERMS FROM THE STRAIT OF MAGELLAN, CHILE 2006 Mutschke RíosDocumento18 páginasSPATIAL DISTRIBUTION AND RELATIVE ABUNDANCE OF ECHINODERMS FROM THE STRAIT OF MAGELLAN, CHILE 2006 Mutschke RíosCatalina Merino YunnissiAún no hay calificaciones
- Castillo-Figueroa (2018) Beyond Specimens: Linking Biological Collections, Functional Ecology and Biodiversity ConservationDocumento6 páginasCastillo-Figueroa (2018) Beyond Specimens: Linking Biological Collections, Functional Ecology and Biodiversity ConservationCatalina Merino YunnissiAún no hay calificaciones
- Clasificacion Manejo y Conservacion de Colecciones Biologicas Una Mirada A Su Importancia para La BiodiversidadDocumento11 páginasClasificacion Manejo y Conservacion de Colecciones Biologicas Una Mirada A Su Importancia para La BiodiversidadCatalina Merino YunnissiAún no hay calificaciones
- Una Mirada Al Mundo de Las Colecciones BiológicasDocumento8 páginasUna Mirada Al Mundo de Las Colecciones BiológicasCatalina Merino YunnissiAún no hay calificaciones
- ProyectoDocumento11 páginasProyectoCatalina Merino YunnissiAún no hay calificaciones
- Documentación ColeccionesDocumento55 páginasDocumentación ColeccionesCatalina Merino YunnissiAún no hay calificaciones
- Herbarios2008Granada GBIF-FPDocumento38 páginasHerbarios2008Granada GBIF-FPCatalina Merino YunnissiAún no hay calificaciones
- Coleccion de Insectos - ECOSURDocumento44 páginasColeccion de Insectos - ECOSURCatalina Merino YunnissiAún no hay calificaciones
- Foro: Sostenibilidad de Los Museos de Historia NaturalDocumento28 páginasForo: Sostenibilidad de Los Museos de Historia NaturalCatalina Merino YunnissiAún no hay calificaciones
- Gestión del Herbario EIF en la Universidad de ChileDocumento29 páginasGestión del Herbario EIF en la Universidad de ChileCatalina Merino YunnissiAún no hay calificaciones
- Rocky Reef Benthic Assemblages in The Magellan Strait and The South Shetland Islands (Antarctica) Newcombe & Cárdenas 2011Documento12 páginasRocky Reef Benthic Assemblages in The Magellan Strait and The South Shetland Islands (Antarctica) Newcombe & Cárdenas 2011Catalina Merino YunnissiAún no hay calificaciones
- Semaforizacion Reactivos Lab. Clinico 2016Documento11 páginasSemaforizacion Reactivos Lab. Clinico 2016Laura ParraAún no hay calificaciones
- Riesgo Ergonómico, Higiene Postural y Pausas ActivasDocumento26 páginasRiesgo Ergonómico, Higiene Postural y Pausas ActivasJhon Fredy RamírezAún no hay calificaciones
- Linfoma de HodgkinDocumento20 páginasLinfoma de HodgkinMarianella Saravia LopezAún no hay calificaciones
- Cómo Corregir Errores de PronunciaciónDocumento10 páginasCómo Corregir Errores de PronunciaciónOdette Rivera GalleguillosAún no hay calificaciones
- Ética AmbientalDocumento16 páginasÉtica AmbientalNancyAún no hay calificaciones
- Reporte de Practica Aparatos ReproductoresDocumento9 páginasReporte de Practica Aparatos ReproductoresRoberto BelloAún no hay calificaciones
- Cartilla Producción AgricolaDocumento84 páginasCartilla Producción AgricolaakariAún no hay calificaciones
- Técnicas de coproparasitología para el análisis macroscópico y microscópico de hecesDocumento8 páginasTécnicas de coproparasitología para el análisis macroscópico y microscópico de hecesOmar Vega RuizAún no hay calificaciones
- Ritmos BiológicosDocumento9 páginasRitmos BiológicosMoises Escobar Merlin100% (1)
- Infografia 2 - 7 REPLICACION, TRANSCRIPCION Y TRADUCCION DEL ADNDocumento3 páginasInfografia 2 - 7 REPLICACION, TRANSCRIPCION Y TRADUCCION DEL ADNYERSON DANILO MORENO HUAROTOAún no hay calificaciones
- Primer Parcial de Anatomia AnatomiaDocumento40 páginasPrimer Parcial de Anatomia AnatomiaMaria De Los Angeles MoraAún no hay calificaciones
- Las 4 Etapas Del Desarrollo Cognitivo de Jean PiagetDocumento4 páginasLas 4 Etapas Del Desarrollo Cognitivo de Jean PiagetPablo Aguilera PachecoAún no hay calificaciones
- TeoriaDocumento6 páginasTeoriaSandra RenteríaAún no hay calificaciones
- Indice de Confort de La VegetacionDocumento22 páginasIndice de Confort de La VegetacionOrlando Jiménez GonzálezAún no hay calificaciones
- 7a Práctica Lab Anatomia Riñón y UréteresDocumento3 páginas7a Práctica Lab Anatomia Riñón y UréteresRuth Cobo RosalesAún no hay calificaciones
- Medicina Interna Renal ResumenDocumento54 páginasMedicina Interna Renal ResumenSharan MalkaniAún no hay calificaciones
- Fichas Bibliograficas de ContenidoDocumento25 páginasFichas Bibliograficas de ContenidoJavier Sánchez CAún no hay calificaciones
- Diabetes Insipida GestacionalDocumento7 páginasDiabetes Insipida Gestacionalingrid QuiñonesAún no hay calificaciones
- EL ABORTO, Estudio BiblicoDocumento8 páginasEL ABORTO, Estudio BiblicoLuisMoraAún no hay calificaciones
- Screening Battelle (1) CompletoDocumento6 páginasScreening Battelle (1) CompletoandreamarinAún no hay calificaciones
- Literatura y Cultura. Una Introducción (Miguel Ángel Huamán)Documento23 páginasLiteratura y Cultura. Una Introducción (Miguel Ángel Huamán)Raul A. Burneo100% (1)
- Razonamiento Verbal 02 Repaso Miscelánea t.tDocumento2 páginasRazonamiento Verbal 02 Repaso Miscelánea t.tmelani18moonAún no hay calificaciones
- Trabajo Nro 3Documento29 páginasTrabajo Nro 3Arven CastilloAún no hay calificaciones
- Soluciones Tampones y Sistemas Tampones Biológicos (San Juan Bautista)Documento14 páginasSoluciones Tampones y Sistemas Tampones Biológicos (San Juan Bautista)Renzo Alfredo Céspedes HerediaAún no hay calificaciones
- Propiedades (2) 1Documento3 páginasPropiedades (2) 1Mariano SilveyraAún no hay calificaciones
- Exposición de CarnesDocumento22 páginasExposición de CarnesJeniferYraricaParedesAún no hay calificaciones
- Masaje EroticoDocumento7 páginasMasaje EroticoAndrea TinocoAún no hay calificaciones
- ConserjeDocumento4 páginasConserjeRodolfo OspìnaAún no hay calificaciones
- Ejercicios Propuesto Cat 2023Documento5 páginasEjercicios Propuesto Cat 2023Andrea Sánchez SalasAún no hay calificaciones
- Produccion de Manzana en El Peru (Grupo2)Documento36 páginasProduccion de Manzana en El Peru (Grupo2)Jonathan Gaspar Mendivil100% (1)