También podría gustarte
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017De EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Aún no hay calificaciones
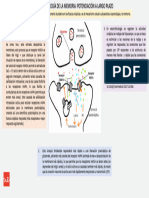
- Potenciación de Largo PlazoDocumento4 páginasPotenciación de Largo PlazoYamila BarberioAún no hay calificaciones
- FisiologiaDocumento32 páginasFisiologiawilliamAún no hay calificaciones
- Tema 6 FisiologicaDocumento7 páginasTema 6 FisiologicaACRAún no hay calificaciones
- Preguntes FISIODocumento8 páginasPreguntes FISIOmargotAún no hay calificaciones
- Potenciación A Largo PlazoDocumento2 páginasPotenciación A Largo PlazoagusalienAún no hay calificaciones
- Potenciación A Largo PlazoDocumento2 páginasPotenciación A Largo PlazoAlexis Cardenas CardenasAún no hay calificaciones
- Tema 3 Infografía Neurofisiología de la memoriaDocumento1 páginaTema 3 Infografía Neurofisiología de la memoriaAlejandroRamírezCapillaAún no hay calificaciones
- TEMA 3 Modulación Neuroquímica de La Memoria PLPDocumento3 páginasTEMA 3 Modulación Neuroquímica de La Memoria PLPFran NistelrooyAún no hay calificaciones
- Las Bases Moleculares de La MemoriaDocumento4 páginasLas Bases Moleculares de La MemoriaCristhian Francis AtamariAún no hay calificaciones
- Sustrato Molecular y Celular de La Plasticidad SinápticaDocumento7 páginasSustrato Molecular y Celular de La Plasticidad SinápticaGeraldine IbacetaAún no hay calificaciones
- TEMA 7. Neurobiologiìa Del Aprendizaje y La MemoriaDocumento39 páginasTEMA 7. Neurobiologiìa Del Aprendizaje y La MemoriaAna100% (1)
- Neuroplasticidad y Plasticidad MuscularDocumento10 páginasNeuroplasticidad y Plasticidad MuscularMacho Peruano Que se RespetaAún no hay calificaciones
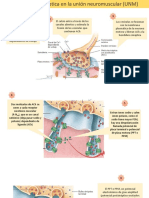
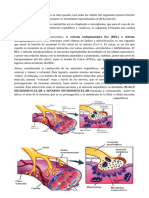
- Union NeuromuscularDocumento3 páginasUnion NeuromuscularMajo FerruscaAún no hay calificaciones
- Fisiología epilepsia: excitación-inhibiciónDocumento3 páginasFisiología epilepsia: excitación-inhibiciónRoxana Ceron SaccacoAún no hay calificaciones
- 1 Neuroplasticidad 3Documento3 páginas1 Neuroplasticidad 3Barbara Bernal RojasAún no hay calificaciones
- Neuroplasticidad (Ficha de Cátedra)Documento6 páginasNeuroplasticidad (Ficha de Cátedra)damianAún no hay calificaciones
- NeurotransmisoresDocumento18 páginasNeurotransmisoresAgustina GuzmánAún no hay calificaciones
- Sistema NitrergicoDocumento1 páginaSistema NitrergicoEleazar Alexander Sánchez Díaz100% (1)
- RESUMEN CAP 7 NEUROBIOLOGÍA DEL APRENDIZAJE Y LA MEMORIA (CURSO 2019-2020)Documento24 páginasRESUMEN CAP 7 NEUROBIOLOGÍA DEL APRENDIZAJE Y LA MEMORIA (CURSO 2019-2020)celiarsjobAún no hay calificaciones
- Tema 7 FisioDocumento12 páginasTema 7 FisioAna Bartual GarciaAún no hay calificaciones
- LA MEMORIA: PROCESOS COGNITIVOS Y NEURONALESDocumento1 páginaLA MEMORIA: PROCESOS COGNITIVOS Y NEURONALESDayanna PalominoAún no hay calificaciones
- Memoria A Medio PlazoDocumento4 páginasMemoria A Medio PlazoZabdi Ochaeta FrutosAún no hay calificaciones
- Fisio Neuronas y Sinapsis NeuronalDocumento5 páginasFisio Neuronas y Sinapsis NeuronalKarina DíazAún no hay calificaciones
- Aumentar la fuerza muscular: sumación espacial y temporalDocumento24 páginasAumentar la fuerza muscular: sumación espacial y temporalmagalimutAún no hay calificaciones
- Atp Contraccion MuscularDocumento5 páginasAtp Contraccion MuscularmonicaAún no hay calificaciones
- TranscripciónDocumento17 páginasTranscripciónroquehernandezmonseAún no hay calificaciones
- Clase 3 Sinapsis ComentariosDocumento32 páginasClase 3 Sinapsis ComentariosSirwinston Churchill Jr.Aún no hay calificaciones
- Work Out SheetDocumento4 páginasWork Out SheetMUS01Aún no hay calificaciones
- Sinapsis ResumenDocumento9 páginasSinapsis ResumenAniretac Neko Seyer50% (4)
- Psicofisiologia Apuntes Unidad 3 y 4Documento57 páginasPsicofisiologia Apuntes Unidad 3 y 4Riely subs;Aún no hay calificaciones
- FISIOPATOLOGIADocumento15 páginasFISIOPATOLOGIAmariaAún no hay calificaciones
- Seminario 4 - MemoriaDocumento16 páginasSeminario 4 - MemoriaLourdesParedesMedinaAún no hay calificaciones
- NeurotransmisoresDocumento72 páginasNeurotransmisoresAyuwoki Hee heeAún no hay calificaciones
- Tema 13 NeurotrasmisoresDocumento7 páginasTema 13 NeurotrasmisoresFrancisco PabonAún no hay calificaciones
- Potencial de Acción, Sinápsis y Neurotransmisores-Guias de Estudio PsicobiologíaDocumento8 páginasPotencial de Acción, Sinápsis y Neurotransmisores-Guias de Estudio PsicobiologíaJohan BallesterosAún no hay calificaciones
- Transmision SinapticaDocumento8 páginasTransmision SinapticaALICIA LISETH VENTURA TINEOAún no hay calificaciones
- Mecanismo de Señalización SinápticasDocumento10 páginasMecanismo de Señalización SinápticasDeborah LopezAún no hay calificaciones
- Asignación SINAPSIS y NEUROTRANSMISORESDocumento7 páginasAsignación SINAPSIS y NEUROTRANSMISORESPaola AlmonteAún no hay calificaciones
- Aprendizaje memoria lenguaje habla corteza cerebral 38Documento10 páginasAprendizaje memoria lenguaje habla corteza cerebral 38Juan David LatinezAún no hay calificaciones
- TS AlglDocumento6 páginasTS AlglBrandon Moscosa MauroAún no hay calificaciones
- Tipos de SinapsisDocumento4 páginasTipos de SinapsisGraciela Carcamo HerreraAún no hay calificaciones
- Tema 4Documento8 páginasTema 4Jaime OrtizAún no hay calificaciones
- Bomba Sodio-PotacioDocumento7 páginasBomba Sodio-PotacioJose Anthony GarciaAún no hay calificaciones
- EXPO FISIO Papel Dela Adrenalina y La Acetilcolina en La SinapsisDocumento5 páginasEXPO FISIO Papel Dela Adrenalina y La Acetilcolina en La SinapsisAlvaro MoralesAún no hay calificaciones
- Rutas de SeñalizaciónDocumento7 páginasRutas de SeñalizaciónFlor de MariaAún no hay calificaciones
- FototransducciónDocumento8 páginasFototransducciónFranyer VargasAún no hay calificaciones
- SINAPSISDocumento35 páginasSINAPSISMiriam Chávez FaustinoAún no hay calificaciones
- Cascadas de SeñalizaciónDocumento2 páginasCascadas de SeñalizaciónAaron CasañasAún no hay calificaciones
- Síndromes de Funciones Mentales SuperioresDocumento25 páginasSíndromes de Funciones Mentales SuperioresAVILA ZEPEDA ARMANDOAún no hay calificaciones
- Mecanismo de La Memoria A Mediano PlazoDocumento4 páginasMecanismo de La Memoria A Mediano Plazoexcadete82% (11)
- Pregunta 3 y 4Documento3 páginasPregunta 3 y 4Astrid TelloAún no hay calificaciones
- Imprimir PsicofisiologíaDocumento8 páginasImprimir PsicofisiologíaIgnacio ChinaskiAún no hay calificaciones
- Musculos 2 y 3Documento10 páginasMusculos 2 y 3Ees 5 Esteban EcheverriaAún no hay calificaciones
- Haga Un Diagrama Del Metabolismo de Los NeurotransmisoresDocumento6 páginasHaga Un Diagrama Del Metabolismo de Los NeurotransmisoresCarlos Jair Herrera CamposAún no hay calificaciones
- Potencial de acción: onda eléctrica neuronalDocumento3 páginasPotencial de acción: onda eléctrica neuronalELMER JOEL REYES DIAZAún no hay calificaciones
- Repaso Tema 2Documento6 páginasRepaso Tema 2Sebastian GonzalezAún no hay calificaciones
- Clase 11 de Agosto Fisio 2Documento11 páginasClase 11 de Agosto Fisio 2Brandon GonzalezAún no hay calificaciones
- AMPcDocumento11 páginasAMPcREGINALDOAún no hay calificaciones
- AF5 Material CompletoDocumento38 páginasAF5 Material CompletoNeli Gonzales85% (41)
- Cuaderno de Estimulos 2Documento99 páginasCuaderno de Estimulos 2FernandaAún no hay calificaciones
- NeurotransmisoresDocumento2 páginasNeurotransmisoresFernandaAún no hay calificaciones
- Cuaderno de ANOTACIÓN 1 PDFDocumento18 páginasCuaderno de ANOTACIÓN 1 PDFJuan José Pabón CastroAún no hay calificaciones
- Las Mujeres Que Aman DemasiadoDocumento18 páginasLas Mujeres Que Aman DemasiadoFernandaAún no hay calificaciones
- ITPA Cuaderno de Estimulos 1Documento115 páginasITPA Cuaderno de Estimulos 1Fernanda100% (1)
- Informe Cuestionario ItpaDocumento3 páginasInforme Cuestionario ItpaFernandaAún no hay calificaciones
- Determinación de EnzimasDocumento95 páginasDeterminación de EnzimasStela MarisAún no hay calificaciones
- Tensmed S82: Instrucciones de UsoDocumento36 páginasTensmed S82: Instrucciones de UsoDiego Pizarro MonroyAún no hay calificaciones
- Farmacia Contenidos MinimosDocumento15 páginasFarmacia Contenidos MinimosEvelynAún no hay calificaciones
- Clases Avanzadas Tarjeta CompletaDocumento7 páginasClases Avanzadas Tarjeta Completabellekie paola glesilAún no hay calificaciones
- Guía de Funcion CelularDocumento10 páginasGuía de Funcion CelularDeivi LachapelAún no hay calificaciones
- FISIOLOGÍA Y PSICOLOGÍA CIENTÍFICADocumento4 páginasFISIOLOGÍA Y PSICOLOGÍA CIENTÍFICALaura Camila Santofimio TovarAún no hay calificaciones
- Laboratorio de Inmunología IDocumento25 páginasLaboratorio de Inmunología ILizandra SzottAún no hay calificaciones
- Cerebro!Documento12 páginasCerebro!Natalia Delgado NavarroAún no hay calificaciones
- Semana 2Documento45 páginasSemana 2Erick GomezAún no hay calificaciones
- 4-4 Alumnos Seminario Agua y Electrolitos 2019 CorregidoDocumento62 páginas4-4 Alumnos Seminario Agua y Electrolitos 2019 CorregidoFlor GianoAún no hay calificaciones
- HIDRATOS DE CARBONO Importancia en El MetabolismoDocumento31 páginasHIDRATOS DE CARBONO Importancia en El MetabolismoAdrián ValdiviesoAún no hay calificaciones
- NEUROHIPOFISISDocumento9 páginasNEUROHIPOFISISpaulo cuevaAún no hay calificaciones
- Acuerdos de convivencia en el aulaDocumento27 páginasAcuerdos de convivencia en el aulaAlexandra Guardia SupantaAún no hay calificaciones
- Proceso de Envejecimiento: Cambios Que Se ProducenDocumento15 páginasProceso de Envejecimiento: Cambios Que Se ProducenHeilyn Frias aybarAún no hay calificaciones
- Programa Bioq Basica-2 - 2020Documento4 páginasPrograma Bioq Basica-2 - 2020INGRID ANGELICA GONZALEZ VELASCOAún no hay calificaciones
- Autoestudio 03 IPERC v.F.R.Documento4 páginasAutoestudio 03 IPERC v.F.R.Valeska RomeroAún no hay calificaciones
- Anatomía y fisiología humana Ciencias de la SaludDocumento51 páginasAnatomía y fisiología humana Ciencias de la SaludTania Hernández SánchezAún no hay calificaciones
- Intercambio GaseosoDocumento9 páginasIntercambio GaseosoVero TzinAún no hay calificaciones
- Tejido Conectivo Generalidades V 201710.Documento7 páginasTejido Conectivo Generalidades V 201710.Diana ArevaloAún no hay calificaciones
- TALLER 1 Actualizado REPASO de MÚSCULOS Miembro SuperiorDocumento2 páginasTALLER 1 Actualizado REPASO de MÚSCULOS Miembro SuperiorJanna AvilaAún no hay calificaciones
- OsificaciónDocumento11 páginasOsificaciónAbi MendozaAún no hay calificaciones
- Embriología renalDocumento12 páginasEmbriología renalIngrid Suzana RodriguezAún no hay calificaciones
- Factores Que Influyen Sobre El Proceso de FotosintesisDocumento18 páginasFactores Que Influyen Sobre El Proceso de FotosintesisANGEL DENILSON MONTOYA DAVILAAún no hay calificaciones
- Reino Animalia InvertebradosDocumento5 páginasReino Animalia InvertebradosAna Maria Cullanco GonzalesAún no hay calificaciones
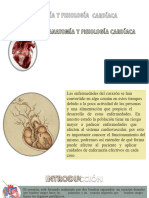
- Anatomia y Fisiologia CardioDocumento20 páginasAnatomia y Fisiologia CardioEnrique EspinoAún no hay calificaciones
- Cuestionario de Músculo CardiacoDocumento3 páginasCuestionario de Músculo CardiacoJesús Leyva GarcíaAún no hay calificaciones
- Act - 06 - Nutrición y Digestion - Ingrid - Maliachi PDFDocumento8 páginasAct - 06 - Nutrición y Digestion - Ingrid - Maliachi PDFIngrid MaliachiAún no hay calificaciones
- RESUMEN Neuro COMPLETO!Documento160 páginasRESUMEN Neuro COMPLETO!aldanaAún no hay calificaciones
- 03 Release Off Balance - Lucia FrancoDocumento482 páginas03 Release Off Balance - Lucia FrancoCésar Ramos Gómez100% (5)
- Cambios Fisicos, Fisiologicos y Psicologicos en El Adulto MayorDocumento23 páginasCambios Fisicos, Fisiologicos y Psicologicos en El Adulto MayorJesus Baldez100% (4)