También podría gustarte
- Captura 2024-02-22 A Las 11.05.36Documento52 páginasCaptura 2024-02-22 A Las 11.05.36marinaAún no hay calificaciones
- Embriologia 13Documento7 páginasEmbriologia 13Duvan Zavaleta BazanAún no hay calificaciones
- Desarrollo de La Primera SemanaDocumento15 páginasDesarrollo de La Primera SemanaFLORENCIA NOELIA DELGADO FERNANDEZAún no hay calificaciones
- Guà - A 3Documento11 páginasGuà - A 3Javier EspinozaAún no hay calificaciones
- Cap 5. GastrulaciónDocumento2 páginasCap 5. Gastrulación4-5 CHARRIS SAMUELAún no hay calificaciones
- Desarrollo Embrion2Documento15 páginasDesarrollo Embrion2Oscar PradoAún no hay calificaciones
- 15-Tema 15 Desarrollo Embrionario GuíaDocumento23 páginas15-Tema 15 Desarrollo Embrionario GuíaPaola MichaelAún no hay calificaciones
- 3º Semana. Gastrulación. Mesodermo y Celoma Intraembrionarios. Neurulación.Documento9 páginas3º Semana. Gastrulación. Mesodermo y Celoma Intraembrionarios. Neurulación.Notnow MaybelaterAún no hay calificaciones
- Embriologia Tema 5. Tercera SemanaDocumento17 páginasEmbriologia Tema 5. Tercera SemanaFranz Quispe CondoriAún no hay calificaciones
- Biologia Del Desarrollo en PecesDocumento3 páginasBiologia Del Desarrollo en PecesSeño Paula Elizabeth GómezAún no hay calificaciones
- Embrio 3Documento9 páginasEmbrio 3Meme Nella CastroAún no hay calificaciones
- Resumen Embriologia Semanas 1 A 8Documento4 páginasResumen Embriologia Semanas 1 A 8Liza Cifuentes100% (1)
- Resumen LangmanDocumento29 páginasResumen Langmanemergencias275200491% (11)
- Wuolah Free Tercera Semana Del Desarrollo EmbrionarioDocumento4 páginasWuolah Free Tercera Semana Del Desarrollo EmbrionariomartintoribiojuliaAún no hay calificaciones
- Etapas Del Desarrollo IntrauterinoDocumento8 páginasEtapas Del Desarrollo IntrauterinoAgostina BugalloAún no hay calificaciones
- DESARROLLO EMBRIOFETAL ResumenDocumento9 páginasDESARROLLO EMBRIOFETAL ResumenJuanAún no hay calificaciones
- Disco Bilaminar y TrilaminarDocumento8 páginasDisco Bilaminar y TrilaminarAnaAún no hay calificaciones
- EmbrioDocumento9 páginasEmbrioYamila Flores Castillo100% (1)
- Núcleo N6Documento22 páginasNúcleo N6Franco TaylorAún no hay calificaciones
- Desarrollo EmbrionarioDocumento3 páginasDesarrollo EmbrionarioThiago VelazquezAún no hay calificaciones
- Em Brio GenesisDocumento28 páginasEm Brio GenesisJOSE RUIZAún no hay calificaciones
- Genes MorfoDocumento34 páginasGenes MorfoJose Luis Maizani LlanosAún no hay calificaciones
- Embriologia de LagmanDocumento5 páginasEmbriologia de LagmanMaria MoraAún no hay calificaciones
- 3era Tercera Semana Del Desarrollo Del Disco Germinativo TrilaminarDocumento15 páginas3era Tercera Semana Del Desarrollo Del Disco Germinativo TrilaminarDaniery MachadoAún no hay calificaciones
- Repaso de EmbriologíaDocumento4 páginasRepaso de EmbriologíaMariana HerreraAún no hay calificaciones
- SEMANA 10 Clase 18. Embriología IIDocumento76 páginasSEMANA 10 Clase 18. Embriología IIPathi VargasAún no hay calificaciones
- Embriología Primera SolemneDocumento18 páginasEmbriología Primera SolemneRocio ContrerasAún no hay calificaciones
- Clase 3Documento117 páginasClase 3Rogger c cutipaAún no hay calificaciones
- Gastrulacic3b3n Delimitacic3b3n y Hojas Embrionarias UtisDocumento54 páginasGastrulacic3b3n Delimitacic3b3n y Hojas Embrionarias UtisLimber Paño YujraAún no hay calificaciones
- Tercera Semana EmbrionDocumento5 páginasTercera Semana EmbrionMar MehhAún no hay calificaciones
- Formación CorporalDocumento30 páginasFormación CorporalMiguel Angel Tristan RamirezAún no hay calificaciones
- Informe Embriologia 1 Exposion Grupo 3Documento16 páginasInforme Embriologia 1 Exposion Grupo 3linsey infante foxAún no hay calificaciones
- Embriologia HumanaDocumento82 páginasEmbriologia HumanaCristhian AlfaroAún no hay calificaciones
- Tercera Semana de Desarrollo: Disco Germinal TrilaminarDocumento6 páginasTercera Semana de Desarrollo: Disco Germinal TrilaminarBrenda Itzel Martinez AgustinAún no hay calificaciones
- EmbriologíaDocumento25 páginasEmbriologíavictoria lizamaAún no hay calificaciones
- GastrulacionDocumento14 páginasGastrulacionErickAún no hay calificaciones
- Anomalías CraneofacialesDocumento27 páginasAnomalías CraneofacialesDaniel EncisoAún no hay calificaciones
- Embriología 5 2da y 3ra SemanaDocumento71 páginasEmbriología 5 2da y 3ra SemanaLUCERO STEPHANY VELASQUEZ YAFACAún no hay calificaciones
- AnatomiaDocumento7 páginasAnatomiaMarta MayoralAún no hay calificaciones
- Capas Germinativas.Documento36 páginasCapas Germinativas.Cecilia Sarahi Villalta GrijalvaAún no hay calificaciones
- Gastrulacion y NeurulacionDocumento6 páginasGastrulacion y NeurulacionVale MAún no hay calificaciones
- 8.clase - Tercera Semana - Gastrulación.Documento20 páginas8.clase - Tercera Semana - Gastrulación.Johanna Mejia BermudezAún no hay calificaciones
- Resumen GastrulacionDocumento2 páginasResumen Gastrulacionfireball764100% (2)
- Em Briolo GiaDocumento4 páginasEm Briolo GiaCamila Oyarzo MelladoAún no hay calificaciones
- Embriología TERCERA Y CUARTA SEMANADocumento16 páginasEmbriología TERCERA Y CUARTA SEMANADuver Arboleda100% (1)
- Periodo Somitico IDocumento3 páginasPeriodo Somitico IChistian DavidAún no hay calificaciones
- Clase 3Documento138 páginasClase 3Eddú OcampoAún no hay calificaciones
- 5º La Gastrulación - Disco GerminativoDocumento7 páginas5º La Gastrulación - Disco GerminativoaleAún no hay calificaciones
- 3era Semana Del EmbarazoDocumento2 páginas3era Semana Del EmbarazoJeandely Ventura RodrguezAún no hay calificaciones
- 8-Segunda y Tercera SemanaDocumento7 páginas8-Segunda y Tercera SemanaLuciana RodríguezAún no hay calificaciones
- Célula y EmbriologíaDocumento10 páginasCélula y EmbriologíaAleejandra riusAún no hay calificaciones
- Tercera Semana Del DesarrolllDocumento11 páginasTercera Semana Del DesarrolllAnghie Katherine Mejia LedezmaAún no hay calificaciones
- Tarea de Embriología-Práctica 5 PDFDocumento5 páginasTarea de Embriología-Práctica 5 PDFNicole SullcaAún no hay calificaciones
- Semana 3 EmbriologiaDocumento18 páginasSemana 3 EmbriologiaAndrea Valenthine Solano MosqueraAún no hay calificaciones
- Etapas de La FecundaciónDocumento3 páginasEtapas de La FecundaciónGardenia EscobarAún no hay calificaciones
- Segunda Semana EmbrioDocumento4 páginasSegunda Semana EmbrioMili GraAún no hay calificaciones
- Embriologia HumanaDocumento14 páginasEmbriologia Humanagolu94Aún no hay calificaciones
- 1-Desarrollo Ontogenetico Del Sistema NerviosoDocumento6 páginas1-Desarrollo Ontogenetico Del Sistema NerviosoRomi SAún no hay calificaciones
- Sem7 CNDocumento6 páginasSem7 CNFede RamirezAún no hay calificaciones
- Sem6 TeratogénesisDocumento3 páginasSem6 TeratogénesisFede RamirezAún no hay calificaciones
- Sem2 SegmentaciónDocumento3 páginasSem2 SegmentaciónFede RamirezAún no hay calificaciones
- Actividades de Embrio SemanalesDocumento3 páginasActividades de Embrio SemanalesFede RamirezAún no hay calificaciones
- Vocabulario Temático de BiologiaDocumento4 páginasVocabulario Temático de BiologiaLuis angel Santiago PérezAún no hay calificaciones
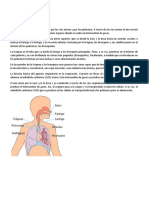
- Aparato RespiratorioDocumento5 páginasAparato RespiratorioLIBRERIAAún no hay calificaciones
- Desarrollo Embrionario Temprano en Vertebrados e InvertebradosDocumento12 páginasDesarrollo Embrionario Temprano en Vertebrados e InvertebradosJuana88% (8)
- Clasificación y Funciones de Las Células Del Sistema InmuneDocumento18 páginasClasificación y Funciones de Las Células Del Sistema Inmunebrenda villaAún no hay calificaciones
- (Cirurgia) Schwartz Principios de Cirugiia 10a EdicionDocumento17 páginas(Cirurgia) Schwartz Principios de Cirugiia 10a EdicionMadmaxooAún no hay calificaciones
- 5° Clase Histo Teoria Apart - Circ.1 UPSJBDocumento29 páginas5° Clase Histo Teoria Apart - Circ.1 UPSJBcielo123456Aún no hay calificaciones
- Guía Practica Laboratorio LuceroDocumento7 páginasGuía Practica Laboratorio LuceroKATHERIN LUCERO FLORES NIEVESAún no hay calificaciones
- GustoDocumento2 páginasGustoAndres Calle HuayancaAún no hay calificaciones
- Sesión de Aprendizaje #04 Ciencia y TecnologíaDocumento7 páginasSesión de Aprendizaje #04 Ciencia y Tecnologíacarmen rosario figueroa roblesAún no hay calificaciones
- Diferecnias Entre Procariota-EucariotaDocumento1 páginaDiferecnias Entre Procariota-Eucariotacesar zavalaAún no hay calificaciones
- Formato Autopsia Medico Legal 1Documento11 páginasFormato Autopsia Medico Legal 1Charli TeheranAún no hay calificaciones
- Músculos Del TóraxDocumento5 páginasMúsculos Del TóraxEstefany MejiaAún no hay calificaciones
- T10. HormonasDocumento8 páginasT10. Hormonasrebeca morata navarroAún no hay calificaciones
- Apuntes de TejidosDocumento41 páginasApuntes de TejidosSol CaraballoAún no hay calificaciones
- El Sistema RespiratorioDocumento9 páginasEl Sistema RespiratorioGrelly IrrazabalAún no hay calificaciones
- Piso PélvicoDocumento41 páginasPiso PélvicoYeni Huamán MejíaAún no hay calificaciones
- Guía 6. Biofísica Respiratoria Clase PrácticaDocumento4 páginasGuía 6. Biofísica Respiratoria Clase PrácticaDiederich JiménezAún no hay calificaciones
- GarcíaLara BOOK ACUPUNCTUREDocumento280 páginasGarcíaLara BOOK ACUPUNCTUREMary CrAún no hay calificaciones
- Prosen CefaloDocumento4 páginasProsen CefaloAitana UribeAún no hay calificaciones
- Conozcamos El Sistema EndocrinoDocumento3 páginasConozcamos El Sistema EndocrinoLAYZA GONZALES FLORAún no hay calificaciones
- RespiratorDocumento5 páginasRespiratorDacostinAún no hay calificaciones
- CAP. 30 Nasal FracturesDocumento16 páginasCAP. 30 Nasal FracturesAn GabrieliAún no hay calificaciones
- Guia 4 - Biologia 6°Documento7 páginasGuia 4 - Biologia 6°JOSE CELESTINO MUTISAún no hay calificaciones
- Isoinmunizacion Materno FetalDocumento5 páginasIsoinmunizacion Materno FetalDANIEL AUDON HERNANDEZ GONZALEZAún no hay calificaciones
- Práctica de Laboratorio 1Documento6 páginasPráctica de Laboratorio 1Nayeli Pongo CarrascoAún no hay calificaciones
- Re Pa SoDocumento21 páginasRe Pa SoFainier Beita MoraAún no hay calificaciones
- Tecnica HistologoDocumento40 páginasTecnica HistologoALESSANDRA LISBETH JUAREZ VIENAAún no hay calificaciones
- Cuestionario de Fisiologia Del Segundo ParcialDocumento5 páginasCuestionario de Fisiologia Del Segundo ParcialRandy LucasAún no hay calificaciones
- Glandula Mamaria PracticaDocumento12 páginasGlandula Mamaria PracticaGerardo Castro RodríguezAún no hay calificaciones
- 2 Reproduccion en El Ser Humano 4 A 8 de MayoDocumento5 páginas2 Reproduccion en El Ser Humano 4 A 8 de MayoLucy DuarteAún no hay calificaciones