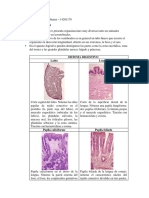
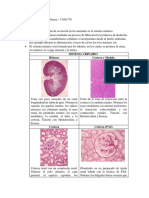
También podría gustarte
- Tema 1 Tecnologia de QuesosDocumento21 páginasTema 1 Tecnologia de QuesosLuz Martinez MartinezAún no hay calificaciones
- Complejo Respiratorio FelinoDocumento38 páginasComplejo Respiratorio FelinoJESUS DAVID FORERO HERNANDEZAún no hay calificaciones
- Propedeutica Veterinaria Z3e Presentacion FinalDocumento19 páginasPropedeutica Veterinaria Z3e Presentacion FinalFernando Cuba rojasAún no hay calificaciones
- Terminología MédicaDocumento25 páginasTerminología MédicaLalo Canalizo Veliz67% (3)
- Desarrollo de Resiliencia - Examen ParcialDocumento4 páginasDesarrollo de Resiliencia - Examen ParcialSofía León GonzalesAún no hay calificaciones
- Bienestar, Enriquecimiento y AdiestramientoDocumento121 páginasBienestar, Enriquecimiento y AdiestramientoAlex ContrerasAún no hay calificaciones
- Nutrición en EquinosDocumento3 páginasNutrición en Equinosalondra martinezAún no hay calificaciones
- Manual de patología clínica en animales de compañíaDe EverandManual de patología clínica en animales de compañíaAún no hay calificaciones
- Protocolo SLC90Documento3 páginasProtocolo SLC90MarcoAún no hay calificaciones
- Oftalmología - I UnidadDocumento5 páginasOftalmología - I UnidadDenis LLontop GuerreroAún no hay calificaciones
- Manual Diagnostico FinalDocumento134 páginasManual Diagnostico FinalSofi Albertini WeilerAún no hay calificaciones
- Manual Anatom A Veterinaria B Sica ParteDocumento129 páginasManual Anatom A Veterinaria B Sica ParteMayra MarroquinAún no hay calificaciones
- Consumo de Carne de CaballoDocumento2 páginasConsumo de Carne de CaballoMANUEL JESÚS HUAMÁN RAMÍREZAún no hay calificaciones
- Anestesia GORDAAADocumento13 páginasAnestesia GORDAAAdaniel enrique jaime lizanaAún no hay calificaciones
- Medidas de Seguridad de Una Vulcanizadora1Documento11 páginasMedidas de Seguridad de Una Vulcanizadora1Jose Zapata RoqueAún no hay calificaciones
- Flut en GatosDocumento14 páginasFlut en Gatosanon_678892511Aún no hay calificaciones
- Bases Moleculares de La HerenciaDocumento79 páginasBases Moleculares de La Herenciastephanie RodriguezAún no hay calificaciones
- Entrevista A PadresDocumento7 páginasEntrevista A PadresLorena Giselle100% (1)
- Manual Fluidoterapia BDocumento42 páginasManual Fluidoterapia BHarold DelgadoAún no hay calificaciones
- Diferencias Anatomicas en AnimalesDocumento262 páginasDiferencias Anatomicas en AnimalesPedro Andrés PupialesAún no hay calificaciones
- Manejo Del Dolor en El Paciente Crítico - Pablo Otero - Mayo2020Documento141 páginasManejo Del Dolor en El Paciente Crítico - Pablo Otero - Mayo2020Dominga MuñozAún no hay calificaciones
- Guía Completa Generalidades Anatomia ULSA 2020 1Documento12 páginasGuía Completa Generalidades Anatomia ULSA 2020 1Laura Sofia Avendaño Morales100% (1)
- Trabajo Del ConejoDocumento16 páginasTrabajo Del ConejoNaleibis NievesAún no hay calificaciones
- Fisiologia Aparato DigestivoDocumento23 páginasFisiologia Aparato DigestivoYURANI VAZQUEZ AGUILERAAún no hay calificaciones
- Complejo Respiratorio FelinoDocumento44 páginasComplejo Respiratorio Felinocheldon jacob100% (1)
- Clase Daño Celular BorboaDocumento31 páginasClase Daño Celular BorboaAlejandro BorboaAún no hay calificaciones
- TGI AvesDocumento13 páginasTGI AvesJuan AndresAún no hay calificaciones
- Fisiologia VeterinariaDocumento15 páginasFisiologia VeterinariaPIZZAAún no hay calificaciones
- MycoplasmaDocumento36 páginasMycoplasmaRosa Bernilla Calderón100% (2)
- Anestesia para Distocia y Anestesia Del Neonato EquinoDocumento9 páginasAnestesia para Distocia y Anestesia Del Neonato EquinoMarjorie SalgadoAún no hay calificaciones
- Sistema Digestivo de CerdosDocumento18 páginasSistema Digestivo de CerdosJosé Lóp'zAún no hay calificaciones
- Medicina Veterinaria UCDocumento2 páginasMedicina Veterinaria UCBryan PalmaAún no hay calificaciones
- Introducción A La Bioquímica-FMVZ-UNA-PunoDocumento17 páginasIntroducción A La Bioquímica-FMVZ-UNA-PunoPedro CoilaAún no hay calificaciones
- Respuestas Cap 1,2,3Documento7 páginasRespuestas Cap 1,2,3Abigail Arandia RosasAún no hay calificaciones
- Los TricotecenosDocumento12 páginasLos TricotecenosChris Tian TerrelAún no hay calificaciones
- Mycobacterium SPPDocumento3 páginasMycobacterium SPPisabelAún no hay calificaciones
- Caso Clinico Cuerpo Extraño Lineal en GatoDocumento4 páginasCaso Clinico Cuerpo Extraño Lineal en GatoferquisAún no hay calificaciones
- Sem 14 Sistema NerviosoDocumento38 páginasSem 14 Sistema NerviosoPiero Noel Salazar HuanascaAún no hay calificaciones
- Anatomia CaninoDocumento10 páginasAnatomia CaninoJosue Enriquez MoralesAún no hay calificaciones
- Tesis 76 Medicina Veterinaria y Zootecnia - CD 454Documento79 páginasTesis 76 Medicina Veterinaria y Zootecnia - CD 454Lesly Huarcaya CondoriAún no hay calificaciones
- Los Analgesicos en Medicina VeterinariaDocumento5 páginasLos Analgesicos en Medicina VeterinariaNancy HernándezAún no hay calificaciones
- Angiología ClasesDocumento23 páginasAngiología ClasesRaul Coila SalazarAún no hay calificaciones
- Anestésicos Inhalatorios VeterinariosDocumento6 páginasAnestésicos Inhalatorios VeterinariosAnitaSchwartzmannBelso100% (1)
- RefractómetroDocumento2 páginasRefractómetroJAIRO SANTIAGO TOVAR UNIGARRO100% (1)
- Manual de Proyecciones RadiograficasDocumento46 páginasManual de Proyecciones RadiograficasLuis Jose Romero OlivaresAún no hay calificaciones
- Sindrome Hemorragico en Cerdos PDFDocumento7 páginasSindrome Hemorragico en Cerdos PDFRicardo Hernandez CruzAún no hay calificaciones
- Guía Anatomia VeterinariaDocumento11 páginasGuía Anatomia VeterinariaChristian Alvarez OrdoñezAún no hay calificaciones
- PRACTICA 1 Exploracion FisicaDocumento10 páginasPRACTICA 1 Exploracion FisicaNoelia Alfonsina Montero VacaAún no hay calificaciones
- Extinción de Los Diversos Tipos de Tortugas en El Estado de Tabasc1Documento6 páginasExtinción de Los Diversos Tipos de Tortugas en El Estado de Tabasc1Karla MartinezAún no hay calificaciones
- 001-2014-CC-Anatomia, Introduccion PDFDocumento40 páginas001-2014-CC-Anatomia, Introduccion PDFgisella0% (1)
- Actividad 1 AnatomiaDocumento11 páginasActividad 1 AnatomiaDavid Garcia SerranoAún no hay calificaciones
- Vademecum 2014 Fasigan Plus o GoldDocumento60 páginasVademecum 2014 Fasigan Plus o GoldAnibal CcasaAún no hay calificaciones
- Digestivo Bovino 2011Documento146 páginasDigestivo Bovino 2011Ainoa Bersani RuizAún no hay calificaciones
- TEXTO BASICO ZOOtDocumento133 páginasTEXTO BASICO ZOOtGabriela Palomo100% (1)
- CASO TYZZER PatologiaDocumento18 páginasCASO TYZZER PatologiaLady CastañedaAún no hay calificaciones
- Manejo de PinguinosDocumento162 páginasManejo de PinguinosChristian Mendoza0% (1)
- Hembra de Shivelly AnatomiaDocumento10 páginasHembra de Shivelly AnatomiaLuis Miguel AmayaAún no hay calificaciones
- Dxnews 112011 PDFDocumento6 páginasDxnews 112011 PDFErnestoRiascosAún no hay calificaciones
- Facultad de Medicina Veterinaria Y Zootecnia: "Sistema NerviosoDocumento29 páginasFacultad de Medicina Veterinaria Y Zootecnia: "Sistema NerviosoLeonel Villaseñor CorteroAún no hay calificaciones
- Abdomen ARP - PpsDocumento106 páginasAbdomen ARP - PpsErnesto RangelAún no hay calificaciones
- Biopsias Veterinarias PDFDocumento5 páginasBiopsias Veterinarias PDFClaudia Azucena Cuellar DiazAún no hay calificaciones
- Tema 1 Citología e Histología VeterinariaDocumento21 páginasTema 1 Citología e Histología VeterinariaCorreoAún no hay calificaciones
- Guía Didáctica Bioquímica I 2017Documento72 páginasGuía Didáctica Bioquímica I 2017ALEJO PS22100% (1)
- PIGMENTOSDocumento59 páginasPIGMENTOSAlejandro BorboaAún no hay calificaciones
- Anon - Plantas MedicinalesDocumento69 páginasAnon - Plantas Medicinalesrory82Aún no hay calificaciones
- Distemper CaninoDocumento71 páginasDistemper Caninohsdaktari61100% (1)
- Manual de prácticas de laboratorio en microbiología veterinariaDe EverandManual de prácticas de laboratorio en microbiología veterinariaAún no hay calificaciones
- Leguminosas espontaneas de posible valor forrajero en ColombiaDe EverandLeguminosas espontaneas de posible valor forrajero en ColombiaAún no hay calificaciones
- Instrumental QuirúrgicoDocumento84 páginasInstrumental Quirúrgicostephanie RodriguezAún no hay calificaciones
- Componentes NutricionalesDocumento5 páginasComponentes Nutricionalesstephanie RodriguezAún no hay calificaciones
- Herencia Mendeliana y Herencia Ligada Al SexoDocumento113 páginasHerencia Mendeliana y Herencia Ligada Al Sexostephanie RodriguezAún no hay calificaciones
- Consumo Voluntario en Mamiferos y Aves RevisionDocumento8 páginasConsumo Voluntario en Mamiferos y Aves Revisionstephanie RodriguezAún no hay calificaciones
- Atlas Osteologia Uab by Nano 1Documento160 páginasAtlas Osteologia Uab by Nano 1Ana GonzálezAún no hay calificaciones
- Preparación Del Campo Operatorio y Personal 2013Documento61 páginasPreparación Del Campo Operatorio y Personal 2013stephanie Rodriguez100% (1)
- Digestivo Semana 1 2022-IDocumento131 páginasDigestivo Semana 1 2022-Istephanie RodriguezAún no hay calificaciones
- Biologia de RobertisDocumento228 páginasBiologia de Robertiskatemu100% (1)
- Bases Moleculares de La MutacionDocumento55 páginasBases Moleculares de La MutacionDiana GonzalezAún no hay calificaciones
- Componentes NutricionalesDocumento5 páginasComponentes Nutricionalesstephanie RodriguezAún no hay calificaciones
- Dominancia y RecesividadDocumento3 páginasDominancia y Recesividadstephanie RodriguezAún no hay calificaciones
- Ontogenia de La ConductaDocumento32 páginasOntogenia de La Conductastephanie RodriguezAún no hay calificaciones
- Comportamiento AnimalDocumento32 páginasComportamiento AnimalSergio Moreno RodriguezAún no hay calificaciones
- TEMA 11. Comportamiento Trófico PDFDocumento27 páginasTEMA 11. Comportamiento Trófico PDFdf710Aún no hay calificaciones
- Componentes NutricionalesDocumento5 páginasComponentes Nutricionalesstephanie RodriguezAún no hay calificaciones
- Componentes NutricionalesDocumento5 páginasComponentes Nutricionalesstephanie RodriguezAún no hay calificaciones
- Atlas Sistema DigestivoDocumento13 páginasAtlas Sistema Digestivostephanie RodriguezAún no hay calificaciones
- Consumo Voluntario en Mamiferos y Aves RevisionDocumento8 páginasConsumo Voluntario en Mamiferos y Aves Revisionstephanie RodriguezAún no hay calificaciones
- 06 07 01 Tema 5Documento32 páginas06 07 01 Tema 5Saymmond CelyAún no hay calificaciones
- Atlas Sistema UrinarioDocumento5 páginasAtlas Sistema Urinariostephanie RodriguezAún no hay calificaciones
- Atlas de HistologiaDocumento16 páginasAtlas de Histologiastephanie RodriguezAún no hay calificaciones
- Atlas de Las Glandulas Anexas Del Sistema DigestivoDocumento5 páginasAtlas de Las Glandulas Anexas Del Sistema Digestivostephanie RodriguezAún no hay calificaciones
- Atlas de HistologiaDocumento16 páginasAtlas de Histologiastephanie RodriguezAún no hay calificaciones
- Atlas Sistema RespiratorioDocumento5 páginasAtlas Sistema Respiratoriostephanie RodriguezAún no hay calificaciones
- CHOQUEDocumento8 páginasCHOQUEMalena S.Aún no hay calificaciones
- Clasificacion ArancelariaDocumento8 páginasClasificacion ArancelariaIsabella Roldán Montaño100% (2)
- PancreatitisDocumento29 páginasPancreatitisBeltor80Aún no hay calificaciones
- Protozoos Tisulares P2Documento5 páginasProtozoos Tisulares P2Noelia Mabel Guzman ClarosAún no hay calificaciones
- Microorganismos LiofilizacionDocumento21 páginasMicroorganismos LiofilizacionLoCV100% (1)
- Detalle de La Visita-2023-02-17-004524410Documento6 páginasDetalle de La Visita-2023-02-17-004524410adela galindoAún no hay calificaciones
- Trabajo Final CoachingDocumento29 páginasTrabajo Final Coachinganny9ariasAún no hay calificaciones
- DISTRIBUCIONESDocumento2 páginasDISTRIBUCIONESBere SánchezAún no hay calificaciones
- Alfonso Longinos Muñoz BenitezDocumento47 páginasAlfonso Longinos Muñoz BenitezMito M GaviriaAún no hay calificaciones
- MELASMASDocumento1 páginaMELASMASVerónica Lévano LudeñaAún no hay calificaciones
- Tab y SAOSDocumento28 páginasTab y SAOSJonathan JimenezAún no hay calificaciones
- PROYECTO FINAL - PROPUESTAS (Evaluación Procesual HITO 5) PDFDocumento2 páginasPROYECTO FINAL - PROPUESTAS (Evaluación Procesual HITO 5) PDFAdrian QuentaAún no hay calificaciones
- Cura para Tdah DietaDocumento2 páginasCura para Tdah DietaVeio MacieiraAún no hay calificaciones
- Via Parenteral (ID, SC, EV, IM)Documento5 páginasVia Parenteral (ID, SC, EV, IM)Adriana Velardez100% (2)
- Cuestionario 8 GametogenesisDocumento2 páginasCuestionario 8 GametogenesisAnnieAún no hay calificaciones
- Proyecto InstructorDocumento11 páginasProyecto InstructorFernando BurgoaAún no hay calificaciones
- Plan Covid-19 Convenio 026-2020-PunDocumento60 páginasPlan Covid-19 Convenio 026-2020-PunHuguier Sánchez Del CastilloAún no hay calificaciones
- Modulo V-Vi - Vii Pasivos Ambientales PDFDocumento69 páginasModulo V-Vi - Vii Pasivos Ambientales PDFPatricio Rodriguez PonteAún no hay calificaciones
- Directorio Violencia C3 (1) Version Final 27 (1) .10.10Documento15 páginasDirectorio Violencia C3 (1) Version Final 27 (1) .10.10api-40881314Aún no hay calificaciones
- SCL-90-R, Iip-64Documento7 páginasSCL-90-R, Iip-64Monica CuellarAún no hay calificaciones
- Terapias de La Tercer GeneracionDocumento10 páginasTerapias de La Tercer GeneracionGlen Moya100% (1)
- Elaboracion de Snacks de OcaDocumento4 páginasElaboracion de Snacks de OcaMAYRA PAMELA100% (1)
- Ensayo Sobre Importancia de Las Emociones en Situacion de Crisis o AdversidadDocumento3 páginasEnsayo Sobre Importancia de Las Emociones en Situacion de Crisis o AdversidadALBA ROCIO CCAHUANA CAÑARIAún no hay calificaciones
- Caso Clinico N°2 HpaDocumento15 páginasCaso Clinico N°2 HpaMilagros Jasmin Julca JulcaAún no hay calificaciones