También podría gustarte
- Emision Forzada de LuzDocumento2 páginasEmision Forzada de LuzRonny GongoraAún no hay calificaciones
- Determinación de Ácido Ascórbico en Jugo Comercial Por Espectroscopia UltravioletaDocumento5 páginasDeterminación de Ácido Ascórbico en Jugo Comercial Por Espectroscopia Ultravioletajaycalderon777Aún no hay calificaciones
- P2 Fluorometr ADocumento5 páginasP2 Fluorometr ADaniel SalazarAún no hay calificaciones
- Circuito LCDocumento1 páginaCircuito LCFranklin F. Fernandez G.Aún no hay calificaciones
- Circuitos de primer ordenDocumento3 páginasCircuitos de primer ordenDanilo FernandezAún no hay calificaciones
- Clase 5 Spin Nuclear y MagnetizaciónDocumento21 páginasClase 5 Spin Nuclear y MagnetizaciónSabina DepierroAún no hay calificaciones
- Determinación del pKa* del primer estado singlete excitado del 2-naftol mediante espectroscopía de fluorescenciaDocumento18 páginasDeterminación del pKa* del primer estado singlete excitado del 2-naftol mediante espectroscopía de fluorescenciaCarlos GiorgiAún no hay calificaciones
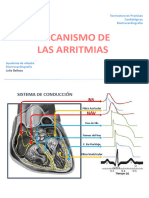
- Mecanismo de ArritmiasDocumento21 páginasMecanismo de Arritmiasyapau14Aún no hay calificaciones
- Flourimetría 2018. Notas Dr. Peón PDFDocumento26 páginasFlourimetría 2018. Notas Dr. Peón PDFManuel LimonAún no hay calificaciones
- Química Clínica 1991 10 (5) 347-359 Espectroscopia DerivadaDocumento13 páginasQuímica Clínica 1991 10 (5) 347-359 Espectroscopia DerivadaFelipe JiménezAún no hay calificaciones
- La Emisión EstimuladaDocumento8 páginasLa Emisión EstimuladaPablo Andres Flores TovarAún no hay calificaciones
- Resonancia ElectricaDocumento7 páginasResonancia ElectricaGabriel BarronAún no hay calificaciones
- Métodos de fluorescencia para análisis químicoDocumento11 páginasMétodos de fluorescencia para análisis químicoalara-731450Aún no hay calificaciones
- Instituto Tecnológico de Piedras Negras: Química e Ingeniería MecatrónicaDocumento2 páginasInstituto Tecnológico de Piedras Negras: Química e Ingeniería MecatrónicaRelvy González LópezAún no hay calificaciones
- Láser de Punto CuánticoDocumento5 páginasLáser de Punto CuánticoManu Santiago HernandezAún no hay calificaciones
- Informe Lab 11 Semana 14Documento25 páginasInforme Lab 11 Semana 14Yunobi Luis Moreno esquivelAún no hay calificaciones
- Circuito ResonanteDocumento2 páginasCircuito ResonantefaustoAún no hay calificaciones
- Clases - Fisiología CardiovascularDocumento76 páginasClases - Fisiología CardiovascularEstefania Gonzalez100% (1)
- Info 5Documento5 páginasInfo 5Gaus Gonzales SáenzAún no hay calificaciones
- Actividad 1Documento6 páginasActividad 1LITZY VALLE RAMIREZAún no hay calificaciones
- ELECTROFISIOLOGIADocumento2 páginasELECTROFISIOLOGIANatalia Benavides CruzAún no hay calificaciones
- Circuito LC Sin PerdidasDocumento3 páginasCircuito LC Sin PerdidasFernando Romero67% (3)
- Apuntes FluorescenciaDocumento6 páginasApuntes FluorescenciaAlexiaSeleniaAún no hay calificaciones
- Métodos ópticos en química analítica: Radiación electromagnética y espectrofotometríaDocumento20 páginasMétodos ópticos en química analítica: Radiación electromagnética y espectrofotometríaluis angelAún no hay calificaciones
- "Espectrometría de Luminiscencia Molecular, Absorcion en El Infrarojo Espestroscopia RamanDocumento29 páginas"Espectrometría de Luminiscencia Molecular, Absorcion en El Infrarojo Espestroscopia RamanMARTIN GILARE QUISPEAún no hay calificaciones
- Experimento de Laboratorio Con NERVEDocumento16 páginasExperimento de Laboratorio Con NERVENicoll Perez ManosalvaAún no hay calificaciones
- Anteproyecto de Sulfato de QuininaDocumento29 páginasAnteproyecto de Sulfato de QuininaMario R Hdz100% (1)
- Circuito RCDocumento1 páginaCircuito RCBeatriz SalazarAún no hay calificaciones
- Proyecto Sensores - IeeeDocumento4 páginasProyecto Sensores - IeeeANDRES FELIPE PEREZ FLORIANAún no hay calificaciones
- Determinación de Quinina Por FluorescenciaDocumento4 páginasDeterminación de Quinina Por FluorescenciaChurrumino Rojas LunaAún no hay calificaciones
- Introduccion A Los Metodos Opticos de AnalisisDocumento57 páginasIntroduccion A Los Metodos Opticos de AnalisisP�REZ AGUILAR ANETTEAún no hay calificaciones
- Informe III Lab Circuitos IIDocumento5 páginasInforme III Lab Circuitos IINevar Arnovi Lopez RoseroAún no hay calificaciones
- Teoria de Circuitos ResonantesDocumento30 páginasTeoria de Circuitos ResonantesMoises DanielAún no hay calificaciones
- Informe 5Documento7 páginasInforme 5Felipe ChaconAún no hay calificaciones
- Resonancia ElectricaDocumento3 páginasResonancia ElectricaMolinaHenry92Aún no hay calificaciones
- Caracterización de las oscilaciones Rabi en vapor de RbDocumento9 páginasCaracterización de las oscilaciones Rabi en vapor de RbBrayan MartinezAún no hay calificaciones
- Espectroscopia Bol 4 Raman SolucionesDocumento5 páginasEspectroscopia Bol 4 Raman SolucionesAdrián J.Aún no hay calificaciones
- Resonancia Magnetica NuclearDocumento7 páginasResonancia Magnetica Nucleareli lopezAún no hay calificaciones
- Modulacion PDFDocumento7 páginasModulacion PDFStephanie Gabriela Molina MontoyaAún no hay calificaciones
- C6 Introducción Métodos EspectrométricosDocumento31 páginasC6 Introducción Métodos EspectrométricosManuel OreamunoAún no hay calificaciones
- Circuito LCDocumento3 páginasCircuito LCUlises TuctoAún no hay calificaciones
- BIOFISICADocumento12 páginasBIOFISICAkatalina TrujilloAún no hay calificaciones
- Circuitos Electricos - A6 - RVDocumento3 páginasCircuitos Electricos - A6 - RVRaul :vAún no hay calificaciones
- Circuitos Resonantes en Serie y ParaleloDocumento7 páginasCircuitos Resonantes en Serie y ParaleloUnTalCristhianAún no hay calificaciones
- La relajación nuclear en RM: conceptos básicosDocumento17 páginasLa relajación nuclear en RM: conceptos básicosayelenAún no hay calificaciones
- Teorico ElectorfisiologiaDocumento9 páginasTeorico ElectorfisiologiaIaraAún no hay calificaciones
- FluorescenciaDocumento3 páginasFluorescenciaElliÖtt Jimeenez ColOradoAún no hay calificaciones
- Carga y Descarga de Un CapacitorDocumento8 páginasCarga y Descarga de Un CapacitorDaniel Choque LeónAún no hay calificaciones
- Laboratorio No2 ASK: Oscilador análogo ASKDocumento8 páginasLaboratorio No2 ASK: Oscilador análogo ASKJose GuzmanAún no hay calificaciones
- Respuesta Deelementos RLCDocumento1 páginaRespuesta Deelementos RLCluisangelleonchavez9Aún no hay calificaciones
- Tema 4 Obtención de La Imagen de IrmDocumento29 páginasTema 4 Obtención de La Imagen de Irmsilvia perezAún no hay calificaciones
- Previo Labo 1Documento5 páginasPrevio Labo 1Mauro Jesús Zambrana CastroAún no hay calificaciones
- La Resonancia MecánicaDocumento12 páginasLa Resonancia MecánicaSamuelDavidToctoCabanillas100% (1)
- Taller Tema 13Documento5 páginasTaller Tema 13Yeyzon Javier Cruz VeraAún no hay calificaciones
- Práctica 2 Circuitos ResonantesDocumento6 páginasPráctica 2 Circuitos ResonantesDavid Leonardo Higuera CamargoAún no hay calificaciones
- Espectrofotometría de Absorción UV VISDocumento27 páginasEspectrofotometría de Absorción UV VISnatalia gomezAún no hay calificaciones
- Líneas TX.: Tema 3 - Medidas Del Desacople de Una LTDocumento30 páginasLíneas TX.: Tema 3 - Medidas Del Desacople de Una LTJULIAN ANDRES OSORNO CASTROAún no hay calificaciones
- Informe Corriente Alterna IIDocumento5 páginasInforme Corriente Alterna IIDenis Coromoto UzcáteguiAún no hay calificaciones
- Proyecto BiorrefineriasDocumento28 páginasProyecto BiorrefineriasJavier Hernandez SerranoAún no hay calificaciones
- Tema 3-Metodo QSARDocumento37 páginasTema 3-Metodo QSARJavier Hernandez SerranoAún no hay calificaciones
- Tema 3 - Parte I - Ingeniería de DisolventesDocumento34 páginasTema 3 - Parte I - Ingeniería de DisolventesJavier Hernandez SerranoAún no hay calificaciones
- Microalgas Cultivo y AplicacionesDocumento205 páginasMicroalgas Cultivo y AplicacionesRubén CalvoAún no hay calificaciones
- Kunak Smart Environment ES WebDocumento56 páginasKunak Smart Environment ES WebJavier Hernandez SerranoAún no hay calificaciones
- Tema 8-4boletín 1 Casos PrácticosDocumento2 páginasTema 8-4boletín 1 Casos PrácticosJavier Hernandez Serrano0% (1)
- Tema 3 - Parte I - Ingeniería de DisolventesDocumento34 páginasTema 3 - Parte I - Ingeniería de DisolventesJavier Hernandez SerranoAún no hay calificaciones
- Gestión ambiental y herramientas de sostenibilidadDocumento76 páginasGestión ambiental y herramientas de sostenibilidadJavier Hernandez SerranoAún no hay calificaciones
- Moreno Grau Et Al. 2020 Junio El Saneamiento en CartagenaDocumento13 páginasMoreno Grau Et Al. 2020 Junio El Saneamiento en CartagenaJavier Hernandez SerranoAún no hay calificaciones
- Las primeras leyes ambientales en la historiaDocumento3 páginasLas primeras leyes ambientales en la historiaJavier Hernandez SerranoAún no hay calificaciones
- RSA - Atenza y Moreno-Grau 2016 1 78-79 802-3369-1-PBDocumento3 páginasRSA - Atenza y Moreno-Grau 2016 1 78-79 802-3369-1-PBJavier Hernandez SerranoAún no hay calificaciones
- Gestión ambiental y herramientas de sostenibilidadDocumento76 páginasGestión ambiental y herramientas de sostenibilidadJavier Hernandez SerranoAún no hay calificaciones
- Herramientas ObligatoriasDocumento8 páginasHerramientas ObligatoriasJavier Hernandez SerranoAún no hay calificaciones
- Herramientas ObligatoriasDocumento8 páginasHerramientas ObligatoriasJavier Hernandez SerranoAún no hay calificaciones
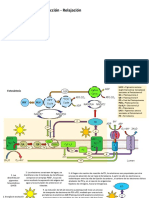
- 1fotosíntesis y Fluorometria ActivaDocumento37 páginas1fotosíntesis y Fluorometria ActivaJavier Hernandez SerranoAún no hay calificaciones
- Moreno Grau Et Al. 2020 Junio El Saneamiento en CartagenaDocumento13 páginasMoreno Grau Et Al. 2020 Junio El Saneamiento en CartagenaJavier Hernandez SerranoAún no hay calificaciones
- MicroalgasDocumento11 páginasMicroalgasLaura Cervera100% (1)
- MicroalgasDocumento11 páginasMicroalgasLaura Cervera100% (1)
- GR17Documento2 páginasGR17Javier Hernandez SerranoAún no hay calificaciones
- 3fundamentos de Fluorometría Activa FIReDocumento30 páginas3fundamentos de Fluorometría Activa FIReJavier Hernandez SerranoAún no hay calificaciones
- 1fotosíntesis y Fluorometria ActivaDocumento37 páginas1fotosíntesis y Fluorometria ActivaJavier Hernandez SerranoAún no hay calificaciones
- GR14Documento4 páginasGR14Javier Hernandez SerranoAún no hay calificaciones
- GR02Documento7 páginasGR02Javier Hernandez SerranoAún no hay calificaciones
- GR16Documento6 páginasGR16Javier Hernandez SerranoAún no hay calificaciones
- GR15Documento4 páginasGR15Javier Hernandez SerranoAún no hay calificaciones
- GR10 PDFDocumento7 páginasGR10 PDFJavier Hernandez SerranoAún no hay calificaciones
- GR18Documento7 páginasGR18Javier Hernandez SerranoAún no hay calificaciones
- Diferencias entre tejido animal y agregados celulares de esponjasDocumento11 páginasDiferencias entre tejido animal y agregados celulares de esponjasJavier Hernandez SerranoAún no hay calificaciones
- Características clave de los cnidariosDocumento6 páginasCaracterísticas clave de los cnidariosJavier Hernandez SerranoAún no hay calificaciones
- Astm en Español 1444Documento19 páginasAstm en Español 1444Antonio SfasAún no hay calificaciones
- Iluminándonos Acerca de Los Códigos ASTM E2297 y E3022: Una Guía para Los Estándares de Lámparas de Luz UV para PNDDocumento6 páginasIluminándonos Acerca de Los Códigos ASTM E2297 y E3022: Una Guía para Los Estándares de Lámparas de Luz UV para PNDmilkamercedesAún no hay calificaciones
- Fosforescencia y FluorescenciaDocumento4 páginasFosforescencia y FluorescenciaJefferson CriolloAún no hay calificaciones
- Histologia Celulas y TejidosDocumento178 páginasHistologia Celulas y Tejidosmaqdv5100% (2)
- Documentologia M1Documento19 páginasDocumentologia M1Mai MuruaAún no hay calificaciones
- FluoresceínaDocumento20 páginasFluoresceínaSebastian GonzalezAún no hay calificaciones
- Tecnica de FluoresenciaDocumento9 páginasTecnica de FluoresenciaFrostye HeladaAún no hay calificaciones
- Manual de Estructura de La Tesis Upav Indice ZeroDocumento25 páginasManual de Estructura de La Tesis Upav Indice ZeroHyde Jekyll YuiAún no hay calificaciones
- Manifestaciones de AceiteDocumento21 páginasManifestaciones de AceiteCristian Ortiz SalazarAún no hay calificaciones
- La Fisica en La CocinaDocumento8 páginasLa Fisica en La CocinainspagintAún no hay calificaciones
- Análisis de falla de equipo: pasos para determinar causaDocumento15 páginasAnálisis de falla de equipo: pasos para determinar causaLuis Alejandro Avila50% (2)
- Histología Básica de Leslie P Gartner y James L. HiattDocumento352 páginasHistología Básica de Leslie P Gartner y James L. Hiattfelipe70% (10)
- Resultados y Analisis de ResuñtadosDocumento2 páginasResultados y Analisis de ResuñtadosvalentinaAún no hay calificaciones
- Líquidos penetrantes y ensayos no destructivosDocumento69 páginasLíquidos penetrantes y ensayos no destructivosLEONCIO RIVERA GALINDOAún no hay calificaciones
- 18 2 Programa de Tecnicas Analiticas InstrumentalesDocumento13 páginas18 2 Programa de Tecnicas Analiticas InstrumentalesClaudiaAún no hay calificaciones
- Rayos X 2015 PDFDocumento33 páginasRayos X 2015 PDFConstanzaGomezAún no hay calificaciones
- ¿Cómo Emite Luz Un Átomo?: El Término Láser Es Un AcróDocumento24 páginas¿Cómo Emite Luz Un Átomo?: El Término Láser Es Un AcróOlga De PilieriAún no hay calificaciones
- Fenómeno de Luminiscencia: Obtención de Fluoresceína (Xanteno) y Luminol (Ftalazina)Documento5 páginasFenómeno de Luminiscencia: Obtención de Fluoresceína (Xanteno) y Luminol (Ftalazina)bazzingaalanAún no hay calificaciones
- Osram Lista de Precios Septiembre 2015Documento52 páginasOsram Lista de Precios Septiembre 2015VEMATELAún no hay calificaciones
- Inspección por líquidos penetrantes: métodos y precaucionesDocumento11 páginasInspección por líquidos penetrantes: métodos y precaucionesGenaro HuarcayaAún no hay calificaciones
- Tipos de Estudios CitometricosDocumento3 páginasTipos de Estudios CitometricosdeinerAún no hay calificaciones
- 2.1 NDT-PT-01Documento38 páginas2.1 NDT-PT-01Carolina PBAún no hay calificaciones
- Espectroscopía de Emisión en FlamaDocumento13 páginasEspectroscopía de Emisión en FlamaNadia AuzsaAún no hay calificaciones
- Detección Raman carotenoidesDocumento16 páginasDetección Raman carotenoidesLeonel AguirreAún no hay calificaciones
- Inspeccion Por Liquidos PenetrantesDocumento12 páginasInspeccion Por Liquidos PenetrantesfilibertoAún no hay calificaciones
- Caracterización de La Materia Orgánica Disuelta en Agua Subterránea Del Valle de Toluca Mediante Espectrofotometría de Fluorescencia 3DDocumento12 páginasCaracterización de La Materia Orgánica Disuelta en Agua Subterránea Del Valle de Toluca Mediante Espectrofotometría de Fluorescencia 3DDAXERAún no hay calificaciones
- Espectroscopía de absorción molecular: Fundamentos y aplicaciones analíticasDocumento49 páginasEspectroscopía de absorción molecular: Fundamentos y aplicaciones analíticasAna MuñozAún no hay calificaciones
- Fotosintesisproductividadyalgasmarinas Eddingetal2006Documento40 páginasFotosintesisproductividadyalgasmarinas Eddingetal2006EgofrutAún no hay calificaciones
- Polvos Reactivos para Revelado de HuellasDocumento5 páginasPolvos Reactivos para Revelado de HuellasSarahí EspinosaAún no hay calificaciones
- Tema 2 - 4 2020Documento127 páginasTema 2 - 4 2020Samuel Antonio Sánchez AlasAún no hay calificaciones