También podría gustarte
- Embriología genital masculina y femeninaDocumento17 páginasEmbriología genital masculina y femeninaAna Cristina Sanchez ZangAún no hay calificaciones
- Desarrollo de Las Gonadas Masculinas y FemeninasDocumento3 páginasDesarrollo de Las Gonadas Masculinas y FemeninasAle AntonioAún no hay calificaciones
- EspermatogenesisDocumento3 páginasEspermatogenesisnumelys mendoza100% (1)
- Embriologã - A Del Aparato Reproductor FemeninoDocumento62 páginasEmbriologã - A Del Aparato Reproductor Femeninojonathan rivas mantillaAún no hay calificaciones
- Desarrollo Embriologico de La Aparato Reproductor FemeninoDocumento41 páginasDesarrollo Embriologico de La Aparato Reproductor FemeninoNoe SotoAún no hay calificaciones
- Gonadas EmbriologiaDocumento1 páginaGonadas EmbriologiaMariana AltamiranoAún no hay calificaciones
- Embriologia (Carlos Lehmann)Documento74 páginasEmbriologia (Carlos Lehmann)AdriánAún no hay calificaciones
- Microbiología II - Sesión 16 - U19 Partes 1 y 2 04.10.2022Documento71 páginasMicrobiología II - Sesión 16 - U19 Partes 1 y 2 04.10.2022felipe carvalhoAún no hay calificaciones
- ParasitologíaDocumento114 páginasParasitologíaTere Mendoza CabezasAún no hay calificaciones
- Anatomia VeterinariaDocumento51 páginasAnatomia VeterinariaCamila AlvarezAún no hay calificaciones
- CestodesDocumento71 páginasCestodesEiliana SantanaAún no hay calificaciones
- Protozoos de Importancia ClínicaDocumento128 páginasProtozoos de Importancia ClínicaДжек ХавокAún no hay calificaciones
- Plexos NerviososDocumento6 páginasPlexos NerviososJosefa OdgersAún no hay calificaciones
- Qué Es La EspermatogénesisDocumento5 páginasQué Es La EspermatogénesisCarola Garrido100% (1)
- Organos genitales femeninos y ciclos reproductoresDocumento27 páginasOrganos genitales femeninos y ciclos reproductoresJackeline ValdiviaAún no hay calificaciones
- Apuntes Reino AnimalDocumento22 páginasApuntes Reino AnimalAna Meiriño MorenoAún no hay calificaciones
- Sistema UrogenitalDocumento7 páginasSistema UrogenitalArturo DíazAún no hay calificaciones
- Aparato Genital Masculino MañanaDocumento9 páginasAparato Genital Masculino MañanaHuamani Flores JhAún no hay calificaciones
- Anatomía y Embriología AnimalDocumento9 páginasAnatomía y Embriología Animalclarimar vasquez cese100% (1)
- Anatomia de Los Organos Sexuales Masculinos y FemeninosDocumento16 páginasAnatomia de Los Organos Sexuales Masculinos y FemeninosJimena Torres Cardenas Stewart100% (1)
- Informe - Practica #4 - IDocumento10 páginasInforme - Practica #4 - ICesar Vargas TacuriAún no hay calificaciones
- Nervios Craneanos Canino PDFDocumento15 páginasNervios Craneanos Canino PDFMarva Carolina Cataldo VargasAún no hay calificaciones
- 7 Aparato ReproductorDocumento17 páginas7 Aparato ReproductorJulio Cesar VegaAún no hay calificaciones
- Espermatogénesis: proceso de formación de espermatozoidesDocumento26 páginasEspermatogénesis: proceso de formación de espermatozoidesEduardo Ocampo100% (1)
- Coccidias ParasitoDocumento73 páginasCoccidias ParasitoLucia VasquezAún no hay calificaciones
- Fosforilacion OxidativaDocumento19 páginasFosforilacion OxidativaQoqe Catalán Durán100% (1)
- Anatomía femenina: órganos reproductivosDocumento5 páginasAnatomía femenina: órganos reproductivosLeidy SaumettAún no hay calificaciones
- OvogénesisDocumento11 páginasOvogénesisMaria Claudia Castro HumpireAún no hay calificaciones
- Meiosis Mitosis y FecundacionDocumento12 páginasMeiosis Mitosis y FecundacionEzequiel Ananias Caceres RojasAún no hay calificaciones
- Parasitologia Oxiuriasis o EnterobiosisDocumento31 páginasParasitologia Oxiuriasis o EnterobiosisAnonymous iisQasXYMSAún no hay calificaciones
- Sistema digestivo del BovinoDocumento54 páginasSistema digestivo del BovinoSANDRA GUTIERREZAún no hay calificaciones
- Seminario N 5Documento16 páginasSeminario N 5Nina Saboya CórdovaAún no hay calificaciones
- Parasitología IiDocumento93 páginasParasitología IiBella Elizabeth CastilloAún no hay calificaciones
- Cromosomas HomólogosDocumento4 páginasCromosomas HomólogosAbelJavierTincoCastroAún no hay calificaciones
- Generalidades de CestodosDocumento41 páginasGeneralidades de CestodosOmar DscAún no hay calificaciones
- 4ta Williams GinecologiaDocumento116 páginas4ta Williams GinecologiaAnthonyArboledaAún no hay calificaciones
- Aparato Reproductor MasculinoDocumento15 páginasAparato Reproductor MasculinoMaritza Lobos HernandezAún no hay calificaciones
- BC-11 & BC-12. Aparato Reproductor Femenino (Héctor Fuenzalida)Documento52 páginasBC-11 & BC-12. Aparato Reproductor Femenino (Héctor Fuenzalida)héctor_wong_5100% (1)
- Desarrollo GonadalDocumento20 páginasDesarrollo GonadalAryadna Bautista MarquezAún no hay calificaciones
- Histología Del OídoDocumento11 páginasHistología Del OídoluisAún no hay calificaciones
- EmbriologíaDocumento17 páginasEmbriologíaJames Russo VelásquezAún no hay calificaciones
- Irrigación de La PelvisDocumento5 páginasIrrigación de La PelvisAzkre KelevraAún no hay calificaciones
- Grupa Muslo 2020Documento43 páginasGrupa Muslo 2020Paola MichaelAún no hay calificaciones
- 4.miologia 99 132Documento34 páginas4.miologia 99 132Pauly BarriosAún no hay calificaciones
- Aparato Reproductor FemeninoDocumento15 páginasAparato Reproductor FemeninoRulas MakAún no hay calificaciones
- OvogenesisDocumento15 páginasOvogenesisRaul CuevaAún no hay calificaciones
- Histologia Sistema EndocrinoDocumento13 páginasHistologia Sistema EndocrinoAndresCajamarcaAún no hay calificaciones
- Céstodos 2 - 2019Documento58 páginasCéstodos 2 - 2019Estefany LopezAún no hay calificaciones
- Transporte de GametosDocumento12 páginasTransporte de GametosPerla MorenoAún no hay calificaciones
- Aparato Rep Macho PDFDocumento44 páginasAparato Rep Macho PDFRo Di IncaAún no hay calificaciones
- GametosDocumento4 páginasGametosGrisel RodriguezAún no hay calificaciones
- ParasitologiaDocumento20 páginasParasitologiajosembvcj4611Aún no hay calificaciones
- 4origen y Diferenciación de GónadasDocumento25 páginas4origen y Diferenciación de GónadasEduardo SandovalAún no hay calificaciones
- 012-2011 AV2-Urogenital Eq y RumDocumento55 páginas012-2011 AV2-Urogenital Eq y RumSandra ChamorroAún no hay calificaciones
- General I Dad EsDocumento51 páginasGeneral I Dad EsJuk CondeAún no hay calificaciones
- Biologia Leyes de Mendel 1203341549314191 3Documento29 páginasBiologia Leyes de Mendel 1203341549314191 3Brayan Andres Guasca CarabaliAún no hay calificaciones
- 005 Miologia Cab, Cue, Tronco PDFDocumento57 páginas005 Miologia Cab, Cue, Tronco PDFDario PalavecinoAún no hay calificaciones
- Embriología y Anatomía Del Aparato Reproductor FemeninoDocumento17 páginasEmbriología y Anatomía Del Aparato Reproductor FemeninoCeci De La Flor CatacoraAún no hay calificaciones
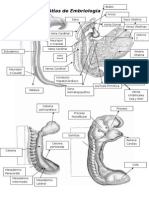
- Atlas de EmbriologíaDocumento4 páginasAtlas de EmbriologíaMedinaAún no hay calificaciones
- Embriologia Sistema Genital PDFDocumento11 páginasEmbriologia Sistema Genital PDFJOCELYN FIERRO GUERREROAún no hay calificaciones
- Formato de Certificado FetalDocumento2 páginasFormato de Certificado FetalDarkinfire WesternAún no hay calificaciones
- Resumen Capitulo 5 LangmanDocumento7 páginasResumen Capitulo 5 LangmanDarkinfire Western100% (1)
- Ácidos GrasosDocumento5 páginasÁcidos GrasosDarkinfire WesternAún no hay calificaciones
- Divisiones de la socio-demografíaDocumento130 páginasDivisiones de la socio-demografíaDarkinfire WesternAún no hay calificaciones
- Defectos renales y del tracto urinarioDocumento2 páginasDefectos renales y del tracto urinarioDarkinfire WesternAún no hay calificaciones
- Capítulo 1 - Introducción A La Regulación y La Señalización MolecularesDocumento2 páginasCapítulo 1 - Introducción A La Regulación y La Señalización MolecularesDarkinfire WesternAún no hay calificaciones
- Aberraciones Cromosómicas en Un Hospital Pediátrico de Tercer Nivel. Anillos de Los Cromosomas 13 y 18Documento20 páginasAberraciones Cromosómicas en Un Hospital Pediátrico de Tercer Nivel. Anillos de Los Cromosomas 13 y 18KylieAún no hay calificaciones
- Produccion de BicarbonatoDocumento12 páginasProduccion de BicarbonatoDarkinfire WesternAún no hay calificaciones
- Plan Prevención de Suicidio - WEB - CASDocumento45 páginasPlan Prevención de Suicidio - WEB - CASPau li100% (3)
- Reabsorcion Del BicarbonatoDocumento12 páginasReabsorcion Del BicarbonatoDarkinfire WesternAún no hay calificaciones
- Apuntes Bioquimica, 19 AgostoDocumento3 páginasApuntes Bioquimica, 19 AgostoDarkinfire WesternAún no hay calificaciones
- Relaciones entre acupuntura y terapia neuralDocumento24 páginasRelaciones entre acupuntura y terapia neuralJose Luis Muela GomezAún no hay calificaciones
- Anatomía y fisiología humana: estructuras y funciones básicasDocumento88 páginasAnatomía y fisiología humana: estructuras y funciones básicasARM ZI4D3Aún no hay calificaciones
- Xilema y floema: los vasos de la circulación vegetalDocumento2 páginasXilema y floema: los vasos de la circulación vegetalximena cerveraAún no hay calificaciones
- TincionesDocumento12 páginasTincionesEmmanuel SerraldeAún no hay calificaciones
- Control 1 de Anatomía y Fisiología BDocumento3 páginasControl 1 de Anatomía y Fisiología BSusiVillaroelLagosAún no hay calificaciones
- Resumen de Educacion FisicaDocumento3 páginasResumen de Educacion FisicaAlexia NavarroAún no hay calificaciones
- Sist CardiovascularDocumento5 páginasSist CardiovascularJohn NeperAún no hay calificaciones
- Celulas Del Tejido ConectivoDocumento2 páginasCelulas Del Tejido Conectivohector manuel coutiño romeroAún no hay calificaciones
- Célula eucariota: estructura y característicasDocumento5 páginasCélula eucariota: estructura y característicasAna ReyesAún no hay calificaciones
- Programa Del Curso de Entrenador Territorial de BalonmanoDocumento20 páginasPrograma Del Curso de Entrenador Territorial de BalonmanoOtsenreAún no hay calificaciones
- Aparato Cardiovascular DMRJDocumento14 páginasAparato Cardiovascular DMRJDanna RamírezAún no hay calificaciones
- Circulación AnimalDocumento5 páginasCirculación AnimalIvan CuenuAún no hay calificaciones
- La PielDocumento31 páginasLa PielAlejandro Colocho100% (1)
- Sistema Circulatorio, Expo BioDocumento1 páginaSistema Circulatorio, Expo BioMuñoz JairAún no hay calificaciones
- Foro N°3 Morfofisiologia Humana: Edinson Aldair Murillo PazDocumento11 páginasForo N°3 Morfofisiologia Humana: Edinson Aldair Murillo PazEDINSON ALDAIR MURILLO PAZAún no hay calificaciones
- ECOE PediDocumento6 páginasECOE PediYomar RiveraAún no hay calificaciones
- 4TO PracticoDocumento3 páginas4TO PracticoFrancisco MaidanaAún no hay calificaciones
- Linea Del Tiempo Desarrollo EmbrionarioDocumento3 páginasLinea Del Tiempo Desarrollo EmbrionarioSalvador Marín100% (2)
- Informe Nº4 Sistema Digestivo en Crustáceos y Moluscos.Documento12 páginasInforme Nº4 Sistema Digestivo en Crustáceos y Moluscos.camila aponte moreanoAún no hay calificaciones
- Anatomia Microscopica de Las Neuronas y Potencial de MembranaDocumento15 páginasAnatomia Microscopica de Las Neuronas y Potencial de MembranaIvan SBAún no hay calificaciones
- Neurología de CabezaDocumento8 páginasNeurología de CabezaJaime Luis Guamán QuintuñaAún no hay calificaciones
- HG 22 4113Documento3 páginasHG 22 4113Margo S CAún no hay calificaciones
- 1.1 La CélulaDocumento18 páginas1.1 La Célulatomas acevedoAún no hay calificaciones
- Calcificaciones Uretra ProstáticaDocumento5 páginasCalcificaciones Uretra ProstáticaRuben Dario Teran DuarteAún no hay calificaciones
- Microbiologia Medica-1 BALZA PDFDocumento177 páginasMicrobiologia Medica-1 BALZA PDFJesus Valdivieso CimmarrustiAún no hay calificaciones
- Formato de Valoracion para FisioterapeutasDocumento14 páginasFormato de Valoracion para FisioterapeutasNikol MolinaAún no hay calificaciones
- El Sistema NerviosoDocumento13 páginasEl Sistema Nervioso04090338Aún no hay calificaciones
- Sistema ExcretorDocumento3 páginasSistema ExcretorDiana Isabel Ochoa YanezAún no hay calificaciones
- PatoDocumento8 páginasPatoDanixaKristelBarriosUcañayAún no hay calificaciones
- Practica Laboratorio 4 Anatomia y Morfologia VegetalDocumento21 páginasPractica Laboratorio 4 Anatomia y Morfologia VegetalFredy MauricioAún no hay calificaciones