También podría gustarte
- Formación Reticular NeuronatomiaDocumento2 páginasFormación Reticular NeuronatomiaHenrySalazarLazoMD100% (4)
- Manual Lab. - CosmetologiaDocumento37 páginasManual Lab. - CosmetologiaGerardo SotoAún no hay calificaciones
- Semana 03 UnasamDocumento51 páginasSemana 03 UnasamWilma Poma RomeroAún no hay calificaciones
- Microbiologia de La CarneeDocumento38 páginasMicrobiologia de La CarneezucoPAún no hay calificaciones
- Horarios de Mantenimiento Del Top Drive TDS-8SADocumento4 páginasHorarios de Mantenimiento Del Top Drive TDS-8SAIrving Antonio Flores AlegriaAún no hay calificaciones
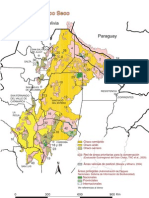
- Chaco SecoDocumento28 páginasChaco SecoHugo CamaraAún no hay calificaciones
- Canales Ionicos Regulados Por VoltajeDocumento24 páginasCanales Ionicos Regulados Por VoltajeAlex Madera100% (1)
- Pozo Tierra Blanca 1113Documento8 páginasPozo Tierra Blanca 1113German DAAún no hay calificaciones
- 12 William Baca Experiencia Peruana Diseno Aplicacion Reforzamiento Estructural Fibra CarbonoDocumento153 páginas12 William Baca Experiencia Peruana Diseno Aplicacion Reforzamiento Estructural Fibra CarbonoEmerson ArteagaAún no hay calificaciones
- Liquidos Corporales 2012Documento38 páginasLiquidos Corporales 2012luisisea100% (1)
- Presentacion A PesquerasDocumento46 páginasPresentacion A PesquerasPedro Miguel SolórzanoAún no hay calificaciones
- Estructura Molecular Del Agua - Tema I - 2014Documento54 páginasEstructura Molecular Del Agua - Tema I - 2014Gregorio Martínez CamposAún no hay calificaciones
- Presentacion Anatomía y Fisiología de L Sistema NerviosoDocumento57 páginasPresentacion Anatomía y Fisiología de L Sistema Nerviosojerson andres sanchez sanchezAún no hay calificaciones
- Aprendizaje de Chaman - Corine SombrumDocumento90 páginasAprendizaje de Chaman - Corine SombrumdommariAún no hay calificaciones
- Manual de AnatomiaDocumento269 páginasManual de AnatomiaDaniel CarbajalAún no hay calificaciones
- La Estructura Molecular Del AguaDocumento13 páginasLa Estructura Molecular Del AguabombmarpAún no hay calificaciones
- Los Saberes Indígenas y La CienciaDocumento25 páginasLos Saberes Indígenas y La CienciaAquiles BailoyóAún no hay calificaciones
- Reporte de Etanol Del VinoDocumento13 páginasReporte de Etanol Del VinoPaola PinedaAún no hay calificaciones
- Riesgo Tomar Ayahuasca Con Personas IncorrectasDocumento7 páginasRiesgo Tomar Ayahuasca Con Personas IncorrectasLucas VillotaAún no hay calificaciones
- La CelulaDocumento28 páginasLa CelulaJoha CarballoAún no hay calificaciones
- P1 Qui OctDocumento6 páginasP1 Qui OctJorge Aranda50% (2)
- 23 Artículo Manuscrito Ensayo 335 2-10-20191121Documento12 páginas23 Artículo Manuscrito Ensayo 335 2-10-20191121Dennys RocioAún no hay calificaciones
- PH de CelulasDocumento10 páginasPH de CelulasStefania Herenandez MotaAún no hay calificaciones
- BacteriasDocumento8 páginasBacteriasJuver HitaloAún no hay calificaciones
- Función de Las EnzimasDocumento5 páginasFunción de Las EnzimasJesus David Garcia CarrilloAún no hay calificaciones
- Manual Del Bio-Tester EMGT PDFDocumento16 páginasManual Del Bio-Tester EMGT PDFLuis Horacio López QuirogaAún no hay calificaciones
- Historia Del MicroscopioDocumento3 páginasHistoria Del MicroscopioÁLèe Aqüiístapáchéé100% (1)
- Radiotrazadores en Medicina NuclearDocumento2 páginasRadiotrazadores en Medicina NuclearAnassinaAún no hay calificaciones
- La Reproducción CelularDocumento30 páginasLa Reproducción CelularAna MariaAún no hay calificaciones
- HistotecnologíaDocumento2 páginasHistotecnologíaMafer LealAún no hay calificaciones
- Division CelularDocumento7 páginasDivision CelularChristian SumariaAún no hay calificaciones
- MicrosDocumento8 páginasMicrosabelAún no hay calificaciones
- Reporte CebollaDocumento5 páginasReporte CebollaFlor ValdizònAún no hay calificaciones
- RADIOTRAZADORESDocumento2 páginasRADIOTRAZADORESAldo Alexander100% (2)
- GENETICADocumento21 páginasGENETICAMelanie N. ToledoAún no hay calificaciones
- AMINOACIDOSDocumento14 páginasAMINOACIDOSestela mendozaAún no hay calificaciones
- Farmaco Sistema NerviosoDocumento91 páginasFarmaco Sistema NerviosoMariela NossaAún no hay calificaciones
- Diversidad CelularDocumento40 páginasDiversidad CelularNiebla001Aún no hay calificaciones
- TRIPLOIDÍADocumento6 páginasTRIPLOIDÍAvelichek67% (3)
- La Division CelularDocumento15 páginasLa Division CelularGustavo Andres Moreno R.100% (2)
- Teoría de La Generación EspontáneaDocumento5 páginasTeoría de La Generación EspontáneaJulio Garcia100% (1)
- El Altar de Muertos Origen y SignificadoDocumento8 páginasEl Altar de Muertos Origen y SignificadoBarrios Estrada Paulina PREPARATORIAAún no hay calificaciones
- Historia de La CélulaDocumento7 páginasHistoria de La CélulaAlender Jesus Vargas TovarAún no hay calificaciones
- Célula EucariotaDocumento8 páginasCélula EucariotaSapmcoolAún no hay calificaciones
- Medio Interno y HomeostasisDocumento3 páginasMedio Interno y Homeostasis54786Aún no hay calificaciones
- Biologica 1 Casi CompletaDocumento60 páginasBiologica 1 Casi CompletaLeonardo FernandezAún no hay calificaciones
- Generalidades de La CelulaDocumento7 páginasGeneralidades de La CelulaAngie Coronado GómezAún no hay calificaciones
- Qué Es Exactamente Un MolDocumento1 páginaQué Es Exactamente Un MolMelanie Rosete CruzAún no hay calificaciones
- CelulaDocumento12 páginasCelulaKalesii Ruiz100% (1)
- Enfermedad de Lyme. Primer Caso Reportado en EcuadorDocumento7 páginasEnfermedad de Lyme. Primer Caso Reportado en EcuadorChristian OrdoñezAún no hay calificaciones
- Diferencias Entre Celulas Procariotas y EucariotasDocumento2 páginasDiferencias Entre Celulas Procariotas y Eucariotasmells19Aún no hay calificaciones
- Adaptaciones Al Frío de Endotermos y EctotermosDocumento3 páginasAdaptaciones Al Frío de Endotermos y EctotermosStefanía Sibille0% (1)
- Fases de La Nucleosíntesis Estelar. Danna Robayo P.Documento4 páginasFases de La Nucleosíntesis Estelar. Danna Robayo P.valentina robayoAún no hay calificaciones
- 1 Neurulación y Creta NeuralDocumento3 páginas1 Neurulación y Creta NeuralDawdunoAún no hay calificaciones
- El Conjunto de Órganos Nerviosos Protegidos Por El Cráneo Constituyen ElDocumento6 páginasEl Conjunto de Órganos Nerviosos Protegidos Por El Cráneo Constituyen ElCristian Valbuena100% (1)
- BiofisicaDocumento25 páginasBiofisicaayelenAún no hay calificaciones
- Plexo BraquialDocumento36 páginasPlexo BraquialYhislein Claudio ZabalaAún no hay calificaciones
- Lectura 1 BQ PDFDocumento4 páginasLectura 1 BQ PDFAlexiss Chavez-De la MoraAún no hay calificaciones
- Fatouh - 3er Ano - Celulas y Estructuras Celulares (Biologia Celular)Documento47 páginasFatouh - 3er Ano - Celulas y Estructuras Celulares (Biologia Celular)api-3824772100% (1)
- Circulacion en PecesDocumento1 páginaCirculacion en PecesMaría Camila Méndez HernándezAún no hay calificaciones
- Respiración Celular y FermentaciónDocumento13 páginasRespiración Celular y FermentaciónCrissRoobinsonAún no hay calificaciones
- Informe de Genetica, MitosisDocumento17 páginasInforme de Genetica, MitosisJesús Darío Carvajal MercadoAún no hay calificaciones
- MitosisDocumento12 páginasMitosisAri VillacrésAún no hay calificaciones
- Activacion Neutrónica InformeDocumento12 páginasActivacion Neutrónica InformeJulio Andre Cabrera CanchariAún no hay calificaciones
- La Molécula de Agua Está Compuesta Por Dos Átomos de Hidrógeno y Uno de Oxígeno Unidos Por Un Enlace CovalenteDocumento6 páginasLa Molécula de Agua Está Compuesta Por Dos Átomos de Hidrógeno y Uno de Oxígeno Unidos Por Un Enlace Covalentemit999666Aún no hay calificaciones
- EL AGUA Y PHDocumento10 páginasEL AGUA Y PHMiranda MontenegroAún no hay calificaciones
- Función Biológica Del Agua..Documento29 páginasFunción Biológica Del Agua..Ricardo Aguado Ornelas0% (2)
- El Agua y La Agua y Estabilidad de Los Alimentos Estabilidad de Los AlimentosDocumento28 páginasEl Agua y La Agua y Estabilidad de Los Alimentos Estabilidad de Los AlimentosSarai GarciaAún no hay calificaciones
- Tintoreria Introduccion 21.10.20Documento28 páginasTintoreria Introduccion 21.10.20MIGUEL RODRIGO CAIRO PAJUELOAún no hay calificaciones
- 1.ficha de Seguridad de Grasas MultiusoDocumento10 páginas1.ficha de Seguridad de Grasas MultiusoLizeth ImpaAún no hay calificaciones
- Arqueobacteria 2Documento25 páginasArqueobacteria 2Roxana VillanuevaAún no hay calificaciones
- PROCESOS DE CONSERVACIÓN Y REDUCCIÓN DE LA MASA (Tecnologia)Documento42 páginasPROCESOS DE CONSERVACIÓN Y REDUCCIÓN DE LA MASA (Tecnologia)Segundo Nieto40% (5)
- Atmofera TerrestreDocumento1 páginaAtmofera TerrestreJesus Juica HuamaniAún no hay calificaciones
- Fosfatos de BayovarDocumento36 páginasFosfatos de BayovarAdrianAún no hay calificaciones
- Disoluciones Acuosas y Equilibrio QuimicoDocumento4 páginasDisoluciones Acuosas y Equilibrio QuimicoCristian Calcina GutierrezAún no hay calificaciones
- Tratamiento Termico de Un AlimentoDocumento3 páginasTratamiento Termico de Un AlimentoEduardo FloresAún no hay calificaciones
- Catalogo PDFDocumento40 páginasCatalogo PDFNathalia Ortiz Serrato0% (1)
- Adrenalina y NoradrenalinaDocumento4 páginasAdrenalina y NoradrenalinaAlejandra RamirezAún no hay calificaciones
- Norma Técnica Peruana 400.022Documento6 páginasNorma Técnica Peruana 400.022Wilmer Tadeo EugenioAún no hay calificaciones
- 03 - Estabilizado GranularDocumento9 páginas03 - Estabilizado Granularpaolo barrionuevoAún no hay calificaciones
- Principio de Le ChatalierDocumento5 páginasPrincipio de Le ChatalierRossman MejicanosAún no hay calificaciones
- Guia 6 Fisica 2Documento4 páginasGuia 6 Fisica 2Ezequiel QuinterosAún no hay calificaciones
- Practica Sintesis de Fenacetina PDFDocumento5 páginasPractica Sintesis de Fenacetina PDFDiana FloresAún no hay calificaciones
- Madera SacDocumento3 páginasMadera SacSara TomateAún no hay calificaciones
- Arequipa Cemento Ip YuraDocumento68 páginasArequipa Cemento Ip YuraRoosevelt Mamani QuispeAún no hay calificaciones
- Metales Alcalinos TerreosDocumento12 páginasMetales Alcalinos TerreosYessica Diaz LimaAún no hay calificaciones
- Ac. Carboxilcos, AmidasDocumento18 páginasAc. Carboxilcos, AmidasJUDITH INGAAún no hay calificaciones
- ES-2179523 T3 FexofenadinaDocumento16 páginasES-2179523 T3 FexofenadinaOscar Eduardo SilvaAún no hay calificaciones
- Aparato RespiratorioDocumento31 páginasAparato Respiratoriolisset8stefany8montoAún no hay calificaciones
- Lab N°1 DETERMINACION DE LA HUMEDAD DEL SUELODocumento5 páginasLab N°1 DETERMINACION DE LA HUMEDAD DEL SUELOCarlos RodriguezAún no hay calificaciones