También podría gustarte
- 5.razas de ConejoDocumento12 páginas5.razas de Conejoana maria depaz barrionuevoAún no hay calificaciones
- AngiospermasDocumento8 páginasAngiospermasMiguel Angel SarmientoAún no hay calificaciones
- Flores ComestiblesDocumento9 páginasFlores ComestiblestomasAún no hay calificaciones
- FloresDocumento35 páginasFloresgatoAún no hay calificaciones
- FlorDocumento15 páginasFlorEduardo CubasAún no hay calificaciones
- FLORDocumento45 páginasFLORJUAN JUNIORAún no hay calificaciones
- FloresDocumento27 páginasFloresMariaTorresAún no hay calificaciones
- Las FloresDocumento20 páginasLas FloresFilial TrujilloAún no hay calificaciones
- FlorDocumento51 páginasFlorMayra CervantesAún no hay calificaciones
- Plantas Espermatofitas Semillas Reproducción Sexual: Nota 1Documento7 páginasPlantas Espermatofitas Semillas Reproducción Sexual: Nota 1MariaTorresAún no hay calificaciones
- FloresDocumento38 páginasFloresAZAún no hay calificaciones
- FlorDocumento38 páginasFlorcristianAún no hay calificaciones
- Las FloresDocumento38 páginasLas FloresMauro Quisbert HuariAún no hay calificaciones
- NubesDocumento14 páginasNubesJuliana OrtizAún no hay calificaciones
- El ABC de La Agricultura Organica y Harina de Rocas Jairo RestrepoDocumento33 páginasEl ABC de La Agricultura Organica y Harina de Rocas Jairo RestrepoAnthony Godoy LoretoAún no hay calificaciones
- FlorDocumento32 páginasFlorJavier Rodríguez MéndezAún no hay calificaciones
- FlorDocumento15 páginasFlorAna Maria Salazar PulgarAún no hay calificaciones
- FloresDocumento67 páginasFloresJonathan Sánchez CastellanosAún no hay calificaciones
- FlorDocumento37 páginasFlorMariaTorresAún no hay calificaciones
- FlorDocumento36 páginasFlorKamila OtaloraAún no hay calificaciones
- FlorDocumento42 páginasFlorMaximo Miranda VeraAún no hay calificaciones
- La FlorDocumento33 páginasLa FlorManuelaAún no hay calificaciones
- FlorDocumento39 páginasFlorrobertAún no hay calificaciones
- FlorDocumento16 páginasFlorkarlaAún no hay calificaciones
- FLORESDocumento3 páginasFLORESJeans Ayquipa RojasAún no hay calificaciones
- FlorDocumento6 páginasFlorelbisAún no hay calificaciones
- Flor - Wikipedia, La Enciclopedia LibreDocumento36 páginasFlor - Wikipedia, La Enciclopedia LibrefoketienefrioAún no hay calificaciones
- La FlorDocumento3 páginasLa FlorRomario Sanchez MendozaAún no hay calificaciones
- FlorDocumento2 páginasFlorJäsmïn CärpïöAún no hay calificaciones
- FlorDocumento10 páginasFlorJanet De la CruzAún no hay calificaciones
- Alma de AmorDocumento3 páginasAlma de Amorjuri ubillusAún no hay calificaciones
- FLORDocumento12 páginasFLORPaulAún no hay calificaciones
- La Flor Es La Estructura Reproductiva Característica de Las Plantas Llamadas Espermatofitas o FanerógamasDocumento5 páginasLa Flor Es La Estructura Reproductiva Característica de Las Plantas Llamadas Espermatofitas o FanerógamasMónica RodriguezAún no hay calificaciones
- FlorDocumento36 páginasFlorgonzalo fernandez de cordobaAún no hay calificaciones
- Botánica IIIDocumento17 páginasBotánica IIIMaxAltairSceneAún no hay calificaciones
- FlorDocumento34 páginasFlorJavierCuaspudBurgosAún no hay calificaciones
- FlorDocumento34 páginasFlorTina SerranoAún no hay calificaciones
- La Flor Es La Estructura Reproductiva Característica de Las Plantas Llamadas Espermatofitas o FanerógamasDocumento2 páginasLa Flor Es La Estructura Reproductiva Característica de Las Plantas Llamadas Espermatofitas o FanerógamasAnita ZuritaAún no hay calificaciones
- Flor EssDocumento249 páginasFlor EssHugo SantanaAún no hay calificaciones
- Definición: Partes de La FlorDocumento2 páginasDefinición: Partes de La Florjuri ubillusAún no hay calificaciones
- La Flor2Documento55 páginasLa Flor2DANIEL MOISES GARCIA ACOSTAAún no hay calificaciones
- FlorDocumento1 páginaFlorFreddy LaricoAún no hay calificaciones
- La Flor Es La Estructura Reproductiva Característica de Las Plantas Llamadas Espermatofitas o FanerógamasDocumento3 páginasLa Flor Es La Estructura Reproductiva Característica de Las Plantas Llamadas Espermatofitas o Fanerógamasjessica pajaroAún no hay calificaciones
- Flores WikiDocumento5 páginasFlores WikiAxel Paul AcostaAún no hay calificaciones
- Apuntes de Gametogénesis, Fecundación y Embriogénesis en AngiospermasDocumento17 páginasApuntes de Gametogénesis, Fecundación y Embriogénesis en AngiospermasJulian y Fernando GilerAún no hay calificaciones
- TP6 Flor e Inflorescencia - BV - LG - 2022Documento30 páginasTP6 Flor e Inflorescencia - BV - LG - 2022Luz CaballeroAún no hay calificaciones
- La Flor IntroduccionDocumento2 páginasLa Flor IntroduccionAymes FernándezAún no hay calificaciones
- FloresDocumento1 páginaFloresJazmany Rolando Mendez NegreteAún no hay calificaciones
- FlorDocumento2 páginasFlorErlyn TorresAún no hay calificaciones
- Cot - KoreanDocumento17 páginasCot - KoreanAndrea EscalanteAún no hay calificaciones
- Estructura Interna y Externa de La FlorDocumento18 páginasEstructura Interna y Externa de La FlorTatiana Doria Perez100% (1)
- Tipos de CitasDocumento2 páginasTipos de CitasLizbeth Mary Velasco CamahualiAún no hay calificaciones
- Texto de Botanica IIDocumento27 páginasTexto de Botanica IISteven Jefry GonzalesAún no hay calificaciones
- FlorDocumento1 páginaFlordianamarcelareyesarellano164Aún no hay calificaciones
- Las FloresDocumento3 páginasLas Floresjose luis andrade mendozaAún no hay calificaciones
- Anatomia - Informe 7Documento18 páginasAnatomia - Informe 7ivanmoran938Aún no hay calificaciones
- Fran Pablo Ramírez Reyes - Colección 3. Estróbilos e InflorescenciasDocumento32 páginasFran Pablo Ramírez Reyes - Colección 3. Estróbilos e InflorescenciasFran RamirezAún no hay calificaciones
- Flor PDFDocumento23 páginasFlor PDFTLvis BenavidesAún no hay calificaciones
- Órganos Vegetales. Flor. Atlas de Histología Vegetal y Animal 8 PDFDocumento9 páginasÓrganos Vegetales. Flor. Atlas de Histología Vegetal y Animal 8 PDFEdwin Enrique Corrales IriarteAún no hay calificaciones
- Flor 2016Documento24 páginasFlor 2016Eduardo YCAún no hay calificaciones
- 1.todas Las RAZAS de GATOS y Sus CaracterísticasDocumento22 páginas1.todas Las RAZAS de GATOS y Sus Característicasana maria depaz barrionuevoAún no hay calificaciones
- 2.cuántos Tipos de Helados ExistenDocumento3 páginas2.cuántos Tipos de Helados Existenana maria depaz barrionuevoAún no hay calificaciones
- 4 HomeDocumento7 páginas4 Homeana maria depaz barrionuevoAún no hay calificaciones
- 3.así Se Llaman Los Distintos Tipos de Zapatos Que Se Venden en Las TiendasDocumento4 páginas3.así Se Llaman Los Distintos Tipos de Zapatos Que Se Venden en Las Tiendasana maria depaz barrionuevoAún no hay calificaciones
- 2 ManzanasDocumento5 páginas2 Manzanasana maria depaz barrionuevoAún no hay calificaciones
- 1.tipos de LimaDocumento9 páginas1.tipos de Limaana maria depaz barrionuevoAún no hay calificaciones
- 4.tipos de AcueductosDocumento2 páginas4.tipos de Acueductosana maria depaz barrionuevoAún no hay calificaciones
- 1.tipos de LimaDocumento9 páginas1.tipos de Limaana maria depaz barrionuevoAún no hay calificaciones
- 5.tipos de JirafasDocumento16 páginas5.tipos de Jirafasana maria depaz barrionuevoAún no hay calificaciones
- 4.tipos de MedidasDocumento2 páginas4.tipos de Medidasana maria depaz barrionuevoAún no hay calificaciones
- 2 ManzanasDocumento5 páginas2 Manzanasana maria depaz barrionuevoAún no hay calificaciones
- 3 MafaldaDocumento6 páginas3 Mafaldaana maria depaz barrionuevoAún no hay calificaciones
- 3 CaptacionesDocumento1 página3 Captacionesana maria depaz barrionuevoAún no hay calificaciones
- 2.tipos de AvesDocumento2 páginas2.tipos de Avesana maria depaz barrionuevoAún no hay calificaciones
- 10 ColorDocumento22 páginas10 Colorana maria depaz barrionuevoAún no hay calificaciones
- Géminis Es Un Signo Mutable Que Forma Parte Del Elemento AireDocumento8 páginasGéminis Es Un Signo Mutable Que Forma Parte Del Elemento Aireana maria depaz barrionuevoAún no hay calificaciones
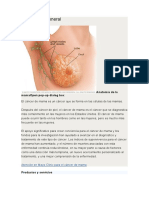
- Cáncer de Cuello UterinoDocumento11 páginasCáncer de Cuello Uterinoana maria depaz barrionuevoAún no hay calificaciones
- 1 PajarosDocumento1 página1 Pajarosana maria depaz barrionuevoAún no hay calificaciones
- Descripción GeneralDocumento13 páginasDescripción Generalana maria depaz barrionuevoAún no hay calificaciones
- DESCRIPCIÓNDocumento1 páginaDESCRIPCIÓNana maria depaz barrionuevoAún no hay calificaciones
- Calaveras TiposDocumento50 páginasCalaveras Tiposana maria depaz barrionuevoAún no hay calificaciones
- 6.perfil de Las ArvejasDocumento1 página6.perfil de Las Arvejasana maria depaz barrionuevoAún no hay calificaciones
- 9.nuevos Nombres para Describir Mejor Los ColoresDocumento7 páginas9.nuevos Nombres para Describir Mejor Los Coloresana maria depaz barrionuevoAún no hay calificaciones
- 8.cultivo de ArvejaDocumento1 página8.cultivo de Arvejaana maria depaz barrionuevoAún no hay calificaciones
- 7.pisum SativumDocumento10 páginas7.pisum Sativumana maria depaz barrionuevoAún no hay calificaciones
- 3.panthera LeoDocumento37 páginas3.panthera Leoana maria depaz barrionuevoAún no hay calificaciones
- 4 VanillaDocumento8 páginas4 Vanillaana maria depaz barrionuevoAún no hay calificaciones
- 5 PandemiaDocumento16 páginas5 Pandemiaana maria depaz barrionuevoAún no hay calificaciones
- 2 LicuadoDocumento1 página2 Licuadoana maria depaz barrionuevoAún no hay calificaciones
- Informe #1Documento9 páginasInforme #1Anonymous DXbZlI4Aún no hay calificaciones
- Unidad Didactica-Las PlantasDocumento3 páginasUnidad Didactica-Las PlantasJess SiCaAún no hay calificaciones
- Cycadophyta PDFDocumento8 páginasCycadophyta PDFReyDemonioAún no hay calificaciones
- 02 Características de Los FrutalesDocumento8 páginas02 Características de Los Frutalesvictorio_elisei100% (1)
- Artes de La PlantaDocumento6 páginasArtes de La PlantagretelAún no hay calificaciones
- .Bulnesia RetamaDocumento1 página.Bulnesia RetamaAgus ButtazzoniAún no hay calificaciones
- OSMedio ArbolesMad PalmerDocumento51 páginasOSMedio ArbolesMad PalmerAntonio De Paz CabelloAún no hay calificaciones
- Biologia Plantas ForrajerasDocumento25 páginasBiologia Plantas ForrajerasRocío Ysabel Sánchez MontoyaAún no hay calificaciones
- 3.2 El Ambiente y Las Plantas - FiogralDocumento39 páginas3.2 El Ambiente y Las Plantas - FiogralZero MadsAún no hay calificaciones
- Maiz Reglas Tecnicas 020721Documento34 páginasMaiz Reglas Tecnicas 020721Bertin MoralesAún no hay calificaciones
- Guía Da Flora Das Fragas Do EumeDocumento108 páginasGuía Da Flora Das Fragas Do EumeJose Manuel MeiraAún no hay calificaciones
- Tema de Las Plantas en InglesDocumento3 páginasTema de Las Plantas en InglesMarién Mateos Fernández100% (1)
- Diseño Randomizado PDFDocumento56 páginasDiseño Randomizado PDFGladys Tanta ChugnasAún no hay calificaciones
- Trabajo Practico N°6 - 2019Documento18 páginasTrabajo Practico N°6 - 2019eliasileAún no hay calificaciones
- Tejido ParenquimaDocumento11 páginasTejido ParenquimaFiona DelfinoAún no hay calificaciones
- Costo de Producción de Arveja ChinaDocumento18 páginasCosto de Producción de Arveja ChinaPedro Benjamin Aguilar GarciaAún no hay calificaciones
- GRUPALDocumento16 páginasGRUPALLina María TarazonaAún no hay calificaciones
- Actividad PlantasDocumento3 páginasActividad PlantasYessii SeclenAún no hay calificaciones
- Introduccion A EspermatofitasDocumento6 páginasIntroduccion A EspermatofitasFredy CamposecoAún no hay calificaciones
- PlantasDocumento5 páginasPlantasJC FernandezAún no hay calificaciones
- Amigo de La NaturalezaDocumento18 páginasAmigo de La Naturalezamaximo lozano0% (1)
- Tecnicas de Recoleccion de Plantas y HerborizacionDocumento9 páginasTecnicas de Recoleccion de Plantas y HerborizacionJohan RodríguezTorresAún no hay calificaciones
- Organografia VegetalDocumento37 páginasOrganografia VegetalWilser DíazAún no hay calificaciones
- Taller Reino Vegetal 1Documento4 páginasTaller Reino Vegetal 1Dylan CRAún no hay calificaciones
- Apio NutricionDocumento23 páginasApio NutricionRoberto Carlos Rojas HuancaAún no hay calificaciones
- Leccion 12 Clasificacion Del Reino VegetalDocumento3 páginasLeccion 12 Clasificacion Del Reino Vegetaldarguin funesAún no hay calificaciones
- Origen y Evolución de Las Angiospermas.Documento6 páginasOrigen y Evolución de Las Angiospermas.Mariangel MariAún no hay calificaciones
- Terreno MorfoooDocumento24 páginasTerreno MorfoooBENJAMIN ZAMORANO ARREDONDOAún no hay calificaciones