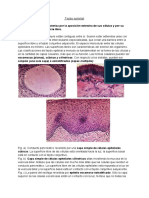
También podría gustarte
- Histologia de Tejidos Epiteliales Terminado CydDocumento17 páginasHistologia de Tejidos Epiteliales Terminado CydSeccion Control de Agencias URIAún no hay calificaciones
- Histología y Embrología del Sitema NerviosoDe EverandHistología y Embrología del Sitema NerviosoCalificación: 4 de 5 estrellas4/5 (4)
- Tecidos Animais 1º Bio Usc PDFDocumento39 páginasTecidos Animais 1º Bio Usc PDFMarta Seco CondeAún no hay calificaciones
- Tarea 2.1 Guia Tejido EpitelialDocumento5 páginasTarea 2.1 Guia Tejido EpitelialMarcheli Nuñez ReyesAún no hay calificaciones
- Tejido EpitelialDocumento12 páginasTejido EpitelialEsaú OrozcoAún no hay calificaciones
- Universidad Autonoma de Baja CaliforniaDocumento11 páginasUniversidad Autonoma de Baja CaliforniaDaniel PerezAún no hay calificaciones
- R 17Documento9 páginasR 17danitzaAún no hay calificaciones
- Generalidades de Los Tejidos BásicosDocumento14 páginasGeneralidades de Los Tejidos BásicosYenifer AriasAún no hay calificaciones
- TEJIDO EPITELIAL VegetalDocumento5 páginasTEJIDO EPITELIAL VegetalcarlosAún no hay calificaciones
- Guia de Estudio de Tejido Epitelial PDFDocumento15 páginasGuia de Estudio de Tejido Epitelial PDFLuisa MatzkAún no hay calificaciones
- CUESTIONARIO DE TEJIDO EPITELIAL InicioDocumento8 páginasCUESTIONARIO DE TEJIDO EPITELIAL InicioLilian Garcia HernándezAún no hay calificaciones
- Tejido EpitelialDocumento4 páginasTejido EpitelialNéstor Alejandro CanelónAún no hay calificaciones
- AnatomíaDocumento4 páginasAnatomíasr.jorgemontano19Aún no hay calificaciones
- Cuestionario Tejido EpitelialDocumento9 páginasCuestionario Tejido EpitelialEnik13Aún no hay calificaciones
- Resumen de Tejido Epitelial y GlandularDocumento14 páginasResumen de Tejido Epitelial y GlandularGenesisAún no hay calificaciones
- TejidosDocumento29 páginasTejidosyqrrkhv2fdAún no hay calificaciones
- Tema 2Documento9 páginasTema 2GLORIA RMAún no hay calificaciones
- TEMA8 Tejidos AnimalDocumento21 páginasTEMA8 Tejidos AnimalClaudia Aldrey GolmarAún no hay calificaciones
- Reyna Gutierrez - Histología - Tejido Epitelial - Unmsm - Facultad de OdontologíaDocumento30 páginasReyna Gutierrez - Histología - Tejido Epitelial - Unmsm - Facultad de OdontologíaGerardo Bardales ArgüellesAún no hay calificaciones
- Comunicación Celular y Epitelios Glandulares David RodriguezDocumento8 páginasComunicación Celular y Epitelios Glandulares David RodriguezDAVID RODRIGUEZ GUERREROAún no hay calificaciones
- Resumen Tejido EpitelialDocumento16 páginasResumen Tejido Epitelialsosa candelaAún no hay calificaciones
- Guía 2, Tejido Epitelial Histo 1 DR Luis Perez MendezDocumento5 páginasGuía 2, Tejido Epitelial Histo 1 DR Luis Perez MendezJuandel Alberto Lalima GomeraAún no hay calificaciones
- Tejido Epitelial y GlandularDocumento23 páginasTejido Epitelial y GlandularGenesisAún no hay calificaciones
- Tejido Epitelial Clase 2Documento4 páginasTejido Epitelial Clase 2Romi CancelarichAún no hay calificaciones
- TejidosDocumento15 páginasTejidosMery MartinezAún no hay calificaciones
- Cuestionario Tejido Epitelial 1 Danna CrisotomoDocumento5 páginasCuestionario Tejido Epitelial 1 Danna CrisotomoDanna michelle Crisotomo quezadaAún no hay calificaciones
- García 4Documento16 páginasGarcía 4Guadalupe GarciaAún no hay calificaciones
- Tema 3Documento4 páginasTema 3elenacanoriveroAún no hay calificaciones
- HISTOLOGÍADocumento12 páginasHISTOLOGÍAnaini honoratoAún no hay calificaciones
- Tejido EpitelialDocumento23 páginasTejido EpitelialJonathan Arizola50% (2)
- CUESTIONARIO TEMA 3 de Morfologia Humana 1 de Lesvia UrbinaDocumento28 páginasCUESTIONARIO TEMA 3 de Morfologia Humana 1 de Lesvia Urbinalesviau44Aún no hay calificaciones
- Capitulo 02 - La Celula y Su Relacion Con El Medio AmbienteDocumento4 páginasCapitulo 02 - La Celula y Su Relacion Con El Medio AmbienteCataIbacetaAún no hay calificaciones
- Tejidos Del Cuerpo HumanoDocumento10 páginasTejidos Del Cuerpo HumanoAngie Lucia Llerena TorresAún no hay calificaciones
- Respuestas Bolillas ERA 1 HISTOLOGÃ - ADocumento25 páginasRespuestas Bolillas ERA 1 HISTOLOGÃ - Amaria trentinAún no hay calificaciones
- Los Epitelios de RevestimientoDocumento2 páginasLos Epitelios de RevestimientoAlejoAún no hay calificaciones
- Tejido EpitelialDocumento4 páginasTejido EpitelialINGRID VALERIA GARCIA REYESAún no hay calificaciones
- TEJIDOSDocumento41 páginasTEJIDOSGerardo PerezAún no hay calificaciones
- Tej Basicos PDFDocumento70 páginasTej Basicos PDFrjds121205Aún no hay calificaciones
- Tejidos AHDocumento51 páginasTejidos AHJHOSIEP GOMEZ SANDOVALAún no hay calificaciones
- Tejido Epitelial PDFDocumento18 páginasTejido Epitelial PDFJennifer RodriguezAún no hay calificaciones
- Biologia 2°Documento36 páginasBiologia 2°luis robinson bocanegra neyraAún no hay calificaciones
- Tejidos Principales Del CuerpoDocumento8 páginasTejidos Principales Del CuerpoMicaela AliAún no hay calificaciones
- EpitelioDocumento3 páginasEpitelioBryan Caja AmanteAún no hay calificaciones
- Tema 2Documento6 páginasTema 2Mireya Garcia NietoAún no hay calificaciones
- TejidosqewdwqerweDocumento18 páginasTejidosqewdwqerweMauricio Nahuel BarrionuevoAún no hay calificaciones
- CUESTIONARIODocumento8 páginasCUESTIONARIOLuciana Cutipa Vásquez MelgarAún no hay calificaciones
- Tejidos Histológicos Unr Medicina FCM 2015Documento45 páginasTejidos Histológicos Unr Medicina FCM 2015Patricia Oroño100% (1)
- Biología Tema 7 Célula Eucariota Estructuras y Orgánulos No MembranososDocumento4 páginasBiología Tema 7 Célula Eucariota Estructuras y Orgánulos No MembranososAnaAún no hay calificaciones
- Copia de Biología 3Documento8 páginasCopia de Biología 3marcoantonio0541Aún no hay calificaciones
- Tejido EpitelialDocumento7 páginasTejido EpitelialLucy SaucedoAún no hay calificaciones
- TEJIDO EPITELIAL 1 ClaseDocumento50 páginasTEJIDO EPITELIAL 1 ClaseEnshi PanicAún no hay calificaciones
- Tejido EpitelialDocumento13 páginasTejido Epitelialtorresriosjuanantonio52Aún no hay calificaciones
- HISTOLOGIADocumento20 páginasHISTOLOGIANerwin MazaAún no hay calificaciones
- Uniones IntercelularesDocumento6 páginasUniones IntercelularesAgustina NuñezAún no hay calificaciones
- Uniones CelularesDocumento22 páginasUniones Celularesjulietttta100% (1)
- Especializaciones de MembranaDocumento6 páginasEspecializaciones de MembranaNany HeddrichAún no hay calificaciones
- Histología Tejido EpitelialDocumento53 páginasHistología Tejido EpitelialLuisa fernanda Chauta MartínezAún no hay calificaciones
- Introducción Tejido EpitelialDocumento3 páginasIntroducción Tejido EpitelialJenny67% (3)
- Apunte 2Documento9 páginasApunte 2Lara ReevesAún no hay calificaciones
- Remuneraciones Unidad 4Documento3 páginasRemuneraciones Unidad 4Soledad RodriguezAún no hay calificaciones
- ESTRUCTURA FUNCIONAL Unidad 2Documento4 páginasESTRUCTURA FUNCIONAL Unidad 2Soledad RodriguezAún no hay calificaciones
- FLUJOGRAMADocumento2 páginasFLUJOGRAMASoledad RodriguezAún no hay calificaciones
- Hogar Del ChivitoDocumento49 páginasHogar Del ChivitoSoledad RodriguezAún no hay calificaciones
- 1 Un Segundo de Coraje Extracto WebDocumento24 páginas1 Un Segundo de Coraje Extracto WebSoledad Rodriguez100% (2)
- Decreto 500Documento51 páginasDecreto 500Soledad RodriguezAún no hay calificaciones
- Crocodylus MoreletiiDocumento4 páginasCrocodylus MoreletiiErick LópezAún no hay calificaciones
- Sistema Tegumentario en Mamíferos.Documento8 páginasSistema Tegumentario en Mamíferos.Selen100% (1)
- Taller de La Clase Circulación en Plantas y Células Grado 4Documento5 páginasTaller de La Clase Circulación en Plantas y Células Grado 4daniel marquez ortegaAún no hay calificaciones
- Pasos para La Diseccion de Un InsectoDocumento1 páginaPasos para La Diseccion de Un Insectoahlcides0% (1)
- Dellman Histologia VeterinariaDocumento206 páginasDellman Histologia VeterinariaAlejandro Barrientos100% (3)
- Esquema InvertebradosDocumento6 páginasEsquema Invertebradoscscrib100% (1)
- 13a Práctica Lab Anatomia de Sistema Reproductor Femenino y MasculinoDocumento3 páginas13a Práctica Lab Anatomia de Sistema Reproductor Femenino y MasculinoRuth Cobo RosalesAún no hay calificaciones
- 2°BASICO - Guia N°6 Ciclo de La Mariposa y El SapoDocumento4 páginas2°BASICO - Guia N°6 Ciclo de La Mariposa y El SapoAlvaro Albano Cid SanchezAún no hay calificaciones
- PrisioneroDocumento101 páginasPrisionerodmone1234100% (1)
- Tipos de PielDocumento8 páginasTipos de Pielsharon perezAún no hay calificaciones
- Antecedentes de La EmbriologiaDocumento3 páginasAntecedentes de La Embriologiaanachirino88% (16)
- Estados Intersexuales Ginecologia Rocio MadridDocumento44 páginasEstados Intersexuales Ginecologia Rocio MadridRocio MadridAún no hay calificaciones
- Echinodermata ChordataDocumento13 páginasEchinodermata ChordataSamuel PrietoAún no hay calificaciones
- Aparato Digestivo Taichi 1Documento23 páginasAparato Digestivo Taichi 1Natalia Moya GallegosAún no hay calificaciones
- Banco de Preguntas - HistoDocumento8 páginasBanco de Preguntas - HistoEduardo R Cano LujánAún no hay calificaciones
- Elaine Marieb - Anatomía y Fisiología Humana9a Ed., Pearson, 2008Documento671 páginasElaine Marieb - Anatomía y Fisiología Humana9a Ed., Pearson, 2008Gabriel Morante100% (1)
- Clase Cestodos PDFDocumento41 páginasClase Cestodos PDFI.E.S.T.P. HIPOLITO UNANUE100% (9)
- Historia de La Anatomía y FisiologíaDocumento4 páginasHistoria de La Anatomía y FisiologíabiologorogerAún no hay calificaciones
- La ReproducciónDocumento34 páginasLa ReproducciónDorisel Soto100% (1)
- Guia de Aprendiz Auntónomo - 2CIENCIAS - SISTEMA CIRCULATORIO Cuarto GradoDocumento3 páginasGuia de Aprendiz Auntónomo - 2CIENCIAS - SISTEMA CIRCULATORIO Cuarto GradodeisyAún no hay calificaciones
- Evaluacion Sistema CirculatorioDocumento2 páginasEvaluacion Sistema CirculatorioAndrea Baquero100% (3)
- Prueba C.naturales Sistema LOcomotor y Nervioso 2016Documento4 páginasPrueba C.naturales Sistema LOcomotor y Nervioso 2016Infeli Herrera MicheaAún no hay calificaciones
- Regeneración (Biología)Documento8 páginasRegeneración (Biología)duannyAún no hay calificaciones
- Tejidos AnimalesDocumento11 páginasTejidos AnimalesErika L Pariasca RojasAún no hay calificaciones
- Médula Ósea y Maduración de LBDocumento109 páginasMédula Ósea y Maduración de LBCasandra Castillo100% (1)
- Sentidos Especiales ETOFDocumento17 páginasSentidos Especiales ETOFMichelle CoronadoAún no hay calificaciones
- Tejido Conjuntivo - Concepto, Tipos y FuncionesDocumento1 páginaTejido Conjuntivo - Concepto, Tipos y FuncionesTania mejia peñaAún no hay calificaciones
- Practica Sistema TegumentarioDocumento28 páginasPractica Sistema TegumentarioSofyChavezAtausinchiAún no hay calificaciones
- Zoolog A I 5to 2024Documento24 páginasZoolog A I 5to 2024josdycornejo17Aún no hay calificaciones
- Sistema NerviosoDocumento15 páginasSistema NerviosoRene Hdez ChAún no hay calificaciones
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- Homo antecessor: El nacimiento de una especieDe EverandHomo antecessor: El nacimiento de una especieCalificación: 5 de 5 estrellas5/5 (1)
- Liberación del trauma: Perdón y temblor es el caminoDe EverandLiberación del trauma: Perdón y temblor es el caminoCalificación: 4 de 5 estrellas4/5 (4)
- 50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaDe Everand50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaCalificación: 4 de 5 estrellas4/5 (37)
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- Cerebro y silencio: Las claves de la creatividad y la serenidadDe EverandCerebro y silencio: Las claves de la creatividad y la serenidadCalificación: 5 de 5 estrellas5/5 (2)
- GuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisDe EverandGuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisCalificación: 4 de 5 estrellas4/5 (9)
- Cuerpo Tóxico: Como Liberar Tu Cuerpo De Las Toxinas Externas E Internas, Y Evitar Asi Los Efectos De Los Radicales LibresDe EverandCuerpo Tóxico: Como Liberar Tu Cuerpo De Las Toxinas Externas E Internas, Y Evitar Asi Los Efectos De Los Radicales LibresCalificación: 5 de 5 estrellas5/5 (2)
- Anatomía del caballo: Guía práctica ilustradaDe EverandAnatomía del caballo: Guía práctica ilustradaCalificación: 4 de 5 estrellas4/5 (9)
- Sesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesDe EverandSesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesCalificación: 4.5 de 5 estrellas4.5/5 (13)
- Psicología forense: Estudio de la mente criminalDe EverandPsicología forense: Estudio de la mente criminalCalificación: 4.5 de 5 estrellas4.5/5 (18)
- Las Enfermedades comienzan y terminan en tu mente: Una guía para la autosanaciónDe EverandLas Enfermedades comienzan y terminan en tu mente: Una guía para la autosanaciónCalificación: 4 de 5 estrellas4/5 (4)
- Contacto & Relación en Psicoterapia: Reflexiones sobre terapia GestaltDe EverandContacto & Relación en Psicoterapia: Reflexiones sobre terapia GestaltCalificación: 3 de 5 estrellas3/5 (2)
- Resumen de Pensar rápido pensar despacio de Daniel KahnemanDe EverandResumen de Pensar rápido pensar despacio de Daniel KahnemanCalificación: 4.5 de 5 estrellas4.5/5 (11)
- Epigenoma: para cuidar tu cuerpo y tu vidaDe EverandEpigenoma: para cuidar tu cuerpo y tu vidaCalificación: 4 de 5 estrellas4/5 (5)
- 200 tareas en terapia breve: 2ª ediciónDe Everand200 tareas en terapia breve: 2ª ediciónCalificación: 4.5 de 5 estrellas4.5/5 (33)
- Las matemáticas de la biología: De las celdas de las abejas a las simetrías de los virusDe EverandLas matemáticas de la biología: De las celdas de las abejas a las simetrías de los virusCalificación: 4 de 5 estrellas4/5 (1)
- Neuroeducación: ¿Cómo aprende el cerebro humano y cómo deberían enseñar los docentes?De EverandNeuroeducación: ¿Cómo aprende el cerebro humano y cómo deberían enseñar los docentes?Calificación: 5 de 5 estrellas5/5 (6)
- Grasas inteligentes: Come más grasa. Pierde más peso. Mantente más sanoDe EverandGrasas inteligentes: Come más grasa. Pierde más peso. Mantente más sanoCalificación: 4 de 5 estrellas4/5 (10)
- El nacimiento del pensamiento científico: Anaximandro de MiletoDe EverandEl nacimiento del pensamiento científico: Anaximandro de MiletoCalificación: 4.5 de 5 estrellas4.5/5 (4)
- UF0085 - Diagnóstico estético facial y corporalDe EverandUF0085 - Diagnóstico estético facial y corporalCalificación: 4 de 5 estrellas4/5 (5)
- Aprender música: ¿Qué nos enseñan las neurociencias del aprendizaje musical?De EverandAprender música: ¿Qué nos enseñan las neurociencias del aprendizaje musical?Calificación: 5 de 5 estrellas5/5 (4)
- Conciencia del tiempo: Por qué pensar como geólogos puede ayudarnos a salvar el planetaDe EverandConciencia del tiempo: Por qué pensar como geólogos puede ayudarnos a salvar el planetaCalificación: 4.5 de 5 estrellas4.5/5 (2)
- El cerebro del niño explicado a los padresDe EverandEl cerebro del niño explicado a los padresCalificación: 4.5 de 5 estrellas4.5/5 (147)