También podría gustarte
- Diversidad de Las InmunoglobulinasDocumento7 páginasDiversidad de Las InmunoglobulinasladbymAún no hay calificaciones
- Estructura y Funcion de Los AnticuerposDocumento3 páginasEstructura y Funcion de Los Anticuerposlucyzoe_2805Aún no hay calificaciones
- Ontogenia de Los Linfocitos T y BDocumento12 páginasOntogenia de Los Linfocitos T y BPaulina Maya100% (1)
- Inmunoglobulina eDocumento26 páginasInmunoglobulina eLpMr Marco AntonioAún no hay calificaciones
- M InmunologiaDocumento126 páginasM InmunologiaHilma Bermudez Gonzalez100% (1)
- 2017 Recombinación de DNADocumento31 páginas2017 Recombinación de DNAMANUELAAún no hay calificaciones
- Resumen Documento Linfocitos BDocumento10 páginasResumen Documento Linfocitos BYesenia Aracely Juarez PopAún no hay calificaciones
- Anticuerpo CREDocumento8 páginasAnticuerpo CRERec 379Aún no hay calificaciones
- INMUNIDocumento14 páginasINMUNIOctavio GarroneAún no hay calificaciones
- Generacion de La Variabilidad de InmunoglobulinasDocumento25 páginasGeneracion de La Variabilidad de InmunoglobulinasMichel Alejandro García100% (1)
- INMUNOGENETICADocumento78 páginasINMUNOGENETICAMiguel VasquezAún no hay calificaciones
- Resumen Clase 5Documento4 páginasResumen Clase 5Dayanna MorenoAún no hay calificaciones
- Tema 3 AnticuerposDocumento6 páginasTema 3 AnticuerposJesus MoralesAún no hay calificaciones
- LB OntogeniaDocumento9 páginasLB OntogeniaBioqui En ProcesoAún no hay calificaciones
- Bloque 4 Preguntas - Adrian Muñoz CastroDocumento6 páginasBloque 4 Preguntas - Adrian Muñoz CastroAdrian MuñozAún no hay calificaciones
- ANTICUERPOSDocumento4 páginasANTICUERPOSOctavio ZmAún no hay calificaciones
- InmunoglobulinasDocumento30 páginasInmunoglobulinasTifoneitorAún no hay calificaciones
- 7 GeneticaigDocumento30 páginas7 GeneticaigElizabeth Román RamirezAún no hay calificaciones
- ESTRUCTURA de Los ANTICUERPOSDocumento7 páginasESTRUCTURA de Los ANTICUERPOSJuan LozadaAún no hay calificaciones
- Anticuerpos Monoclonales y Su Uso Como Tratamiento de Lupus Eritematoso SistémicoDocumento8 páginasAnticuerpos Monoclonales y Su Uso Como Tratamiento de Lupus Eritematoso SistémicoMargarita López AlvarezAún no hay calificaciones
- Kuby Cap 6Documento54 páginasKuby Cap 6cortesmaromba20Aún no hay calificaciones
- Receptores Específicos de Antígeno de Los Linfocitos T y BDocumento8 páginasReceptores Específicos de Antígeno de Los Linfocitos T y BPaulina MayaAún no hay calificaciones
- AnticuerposDocumento15 páginasAnticuerposfrediry david romero aularAún no hay calificaciones
- Aspectos Moleculares y Funcionales de Los AnticuerposDocumento2 páginasAspectos Moleculares y Funcionales de Los AnticuerposDiego Castaño MejiaAún no hay calificaciones
- Organización y Expresión de Los Genes de InmunoglobulinaDocumento78 páginasOrganización y Expresión de Los Genes de InmunoglobulinaSamantha Vazquez100% (1)
- PIA 2 - Grupo 441 - Equipo 6Documento14 páginasPIA 2 - Grupo 441 - Equipo 6rodrigo marroquinAún no hay calificaciones
- Taller Bioquimica 3 AlexaDocumento2 páginasTaller Bioquimica 3 AlexaAlexandra Niño BeltranAún no hay calificaciones
- Mi ParteDocumento2 páginasMi ParteNoa LPAún no hay calificaciones
- Anticuerpos PDFDocumento4 páginasAnticuerpos PDFEmil OCAún no hay calificaciones
- LigandosDocumento7 páginasLigandosNegrita GuevaraAún no hay calificaciones
- Inmunoglobulinas y BCRDocumento14 páginasInmunoglobulinas y BCRLucas Francisco Vargas RodriguezAún no hay calificaciones
- Antigenos y AnticuerposDocumento34 páginasAntigenos y AnticuerposmariaAún no hay calificaciones
- Cuestionario N°3 AnticuerposDocumento5 páginasCuestionario N°3 AnticuerposMaria RadcliffeAún no hay calificaciones
- Clase 8 - Ontogenia BDocumento10 páginasClase 8 - Ontogenia BLautaro VasqueAún no hay calificaciones
- Clase 10. Linfocitos B y Anticuerpos. JHVP. 2017Documento9 páginasClase 10. Linfocitos B y Anticuerpos. JHVP. 2017Yébeny Tufino AlvaradoAún no hay calificaciones
- Triptico de MecaDocumento2 páginasTriptico de MecaAntuanet Geanina Solis CanoAún no hay calificaciones
- Inmunologia T 13Documento6 páginasInmunologia T 13supriyaAún no hay calificaciones
- Apuntes Del Tema de AnticuerposDocumento7 páginasApuntes Del Tema de AnticuerposPaola PintoAún no hay calificaciones
- Anticuerpos o InmunoglobulinasDocumento8 páginasAnticuerpos o InmunoglobulinasL PDAún no hay calificaciones
- INMUNOGLOBULINASDocumento5 páginasINMUNOGLOBULINASLizbthAún no hay calificaciones
- TEMA 5. Anticuerpos y BCRDocumento20 páginasTEMA 5. Anticuerpos y BCRNina Souto BlancoAún no hay calificaciones
- Generación de Diversidad de Linfocitos B y TDocumento23 páginasGeneración de Diversidad de Linfocitos B y TMariano Félix DemasiAún no hay calificaciones
- Tema 9 AnticuerposDocumento40 páginasTema 9 AnticuerposDaniel OrtizAún no hay calificaciones
- Cap 4. Reconocimiento Del Antígeno en El Sistema Inmunitario AdaptativoDocumento11 páginasCap 4. Reconocimiento Del Antígeno en El Sistema Inmunitario AdaptativoSebastián FranciscoAún no hay calificaciones
- Inmuno 1Documento534 páginasInmuno 1Ruth Sinai AguilarAún no hay calificaciones
- S2-1 ANTIGENOS y ANTICUERPOS II UD Inmunología y Serodiagnóstico IIDocumento64 páginasS2-1 ANTIGENOS y ANTICUERPOS II UD Inmunología y Serodiagnóstico IIyesebellAún no hay calificaciones
- Estructura IgGDocumento4 páginasEstructura IgGClaw FollowilAún no hay calificaciones
- Anticuerpos y Respuesta Humoral SecundariaDocumento20 páginasAnticuerpos y Respuesta Humoral SecundariaPabsAún no hay calificaciones
- 05 - Receptores BCR y AnticuerposDocumento4 páginas05 - Receptores BCR y Anticuerpossanti2022Aún no hay calificaciones
- Resumen Del Prmer Parcial de InmunoDocumento19 páginasResumen Del Prmer Parcial de Inmunorocio bogadoAún no hay calificaciones
- AnticuerpoDocumento6 páginasAnticuerpocarolinaAún no hay calificaciones
- Resumen AnticuerposDocumento6 páginasResumen AnticuerposNathalia SerranoAún no hay calificaciones
- Reconocimiento y RespuestaDocumento18 páginasReconocimiento y RespuestaIsabellaAún no hay calificaciones
- InmunoglobulinaDocumento16 páginasInmunoglobulinaVerónica CisnerosAún no hay calificaciones
- Tema 7 - Caracter¡sticas Generales de La L¡nea LinfocitariaDocumento20 páginasTema 7 - Caracter¡sticas Generales de La L¡nea LinfocitariaPablo GarciaAún no hay calificaciones
- In Mu No Globulin AsDocumento10 páginasIn Mu No Globulin AsAndreina LamedaAún no hay calificaciones
- RESUMEN de Inmunoglobulinas M FinalDocumento5 páginasRESUMEN de Inmunoglobulinas M FinalMaría IsabelAún no hay calificaciones
- Receptores de Los LsDocumento11 páginasReceptores de Los LsPaola PintoAún no hay calificaciones
- Cap 7Documento10 páginasCap 7Natalia Espíndola BalderasAún no hay calificaciones
- Taller Division Celular Mitosis y Meiosis - Santiago Tabera LondoñoDocumento7 páginasTaller Division Celular Mitosis y Meiosis - Santiago Tabera LondoñoSantiago LondoñoAún no hay calificaciones
- La Reproducción en Los AnimalesDocumento5 páginasLa Reproducción en Los AnimalesJorge Luis Alva LlajarunaAún no hay calificaciones
- Importancia de La Mitosis y Meiosis YomaDocumento1 páginaImportancia de La Mitosis y Meiosis YomaMakeup YomaAún no hay calificaciones
- Linfocitos T Asesinos Naturales NKTDocumento3 páginasLinfocitos T Asesinos Naturales NKTGaloCoronelAún no hay calificaciones
- Linfocitos T Inmunologia VeterinariaDocumento16 páginasLinfocitos T Inmunologia VeterinariaMel Cajita Feliz :3Aún no hay calificaciones
- Evaluación Mitosis y MeiosisDocumento2 páginasEvaluación Mitosis y MeiosisFacundo Moreno100% (1)
- Reproducción AsexualDocumento3 páginasReproducción AsexualCybernet CoatepequeAún no hay calificaciones
- Celulas MadreDocumento6 páginasCelulas MadreMiranda Lara ItziaAún no hay calificaciones
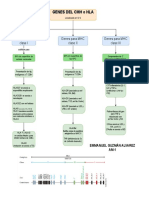
- Genes de HLA o MHCDocumento1 páginaGenes de HLA o MHCEmmanuel GAún no hay calificaciones
- Glosario de EcologíaDocumento2 páginasGlosario de EcologíaItalia CorderoAún no hay calificaciones
- Generalidades Del Sistema InmunitarioDocumento5 páginasGeneralidades Del Sistema InmunitarioMaria Patricia Lopez SantanaAún no hay calificaciones
- Replicación, Transcripción, TraducciónDocumento35 páginasReplicación, Transcripción, Traducciónruth escAún no hay calificaciones
- División CelularDocumento1 páginaDivisión CelularWendy CáceresAún no hay calificaciones
- Reproduccion CelularDocumento2 páginasReproduccion CelularVillca terrazas Dinair JhosetAún no hay calificaciones
- CadenasAlimenticias QuimicaDocumento4 páginasCadenasAlimenticias Quimicasofia tamaraAún no hay calificaciones
- AnticuerpoDocumento6 páginasAnticuerpoLokatikaAún no hay calificaciones
- DOGMA CENTRAL, Transcripción y TraducciónDocumento17 páginasDOGMA CENTRAL, Transcripción y TraducciónK. Lizeth G.CAún no hay calificaciones
- RESPUESTASDocumento1 páginaRESPUESTASCesar TAún no hay calificaciones
- CITOCINASDocumento12 páginasCITOCINASJulio Laurel AriasAún no hay calificaciones
- Mapa Conceptual Disciplinas de La EcologiaDocumento1 páginaMapa Conceptual Disciplinas de La EcologiaLic-emy Atorres100% (2)
- Diccionario Pictórico: Sección: 12-1 Keyci Campos MesénDocumento9 páginasDiccionario Pictórico: Sección: 12-1 Keyci Campos MesénlolAún no hay calificaciones
- Expo Ensayo Diversidad FilogeneticaDocumento9 páginasExpo Ensayo Diversidad Filogeneticaangelica villalob lopezAún no hay calificaciones
- Teoría 2 - Antigenos y AnticuerposDocumento24 páginasTeoría 2 - Antigenos y AnticuerposSharon Almendra de la CruzAún no hay calificaciones
- 7º Guía 22 Ciencias Naturales, ECMDocumento3 páginas7º Guía 22 Ciencias Naturales, ECMIvette Carolina Zelada MartinezAún no hay calificaciones
- 14-Inmunidad Frente A ParásitosDocumento16 páginas14-Inmunidad Frente A Parásitosyccardenasm100% (1)
- Examen EcologíaDocumento2 páginasExamen EcologíaLUIS GONZALO VIDAÑA ESPEJOAún no hay calificaciones
- Generación de Diversidad en Los Linfocitos B yDocumento7 páginasGeneración de Diversidad en Los Linfocitos B yVictormanuel Haro MoriAún no hay calificaciones
- 7° C.NATURALES GUIA DE CLASES No. 2 Flujo de Materia y Energía en Los EcosistemasDocumento7 páginas7° C.NATURALES GUIA DE CLASES No. 2 Flujo de Materia y Energía en Los EcosistemasHeider PadillaAún no hay calificaciones
- Biologia IPNDocumento2 páginasBiologia IPNAdolfoAún no hay calificaciones
- Guía 6 Biología SextoDocumento3 páginasGuía 6 Biología SextoCarmen Carolina CHICAIZA ARANGOAún no hay calificaciones